Citation: Yuanpei Xia, Weisong Zhou, Zhichun Yang. Global analysis and optimal harvesting for a hybrid stochastic phytoplankton-zooplankton-fish model with distributed delays[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 6149-6180. doi: 10.3934/mbe.2020326

| [1] | J. Steele, Stability of Plankton Ecosystem, Chapman and Hall, London, 1974. |
| [2] | T. Saha, M. Bandyopadhyay, Dynamical analysis of toxin producing phytoplankton-zooplankton interactions, Nonlinear Anal. Real World Appl., 10 (2009), 314-332. |
| [3] | Y. Lv, Y. Pei, S. Gao, C. Li, Harvesting of a phytoplankton-zooplankton model, Nonlinear Anal. Real World Appl., 11 (2010), 3608-3619. |
| [4] | C. Liu, L. Wang, Q. Zhang, Y. Yun, Dynamical analysis in a bioeconomic phytoplankton zooplankton system with double time delays and environmental stochasticity, Phys. A, 482 (2017), 682-698. |
| [5] | X. Meng, J. Li, Stability and Hopf bifurcation analysis of a delayed phytoplankton-zooplankton model with Allee effect and linear harvesting, Math. Biosci. Eng., 17 (2019), 1973-2002. |
| [6] | G. Denaro, D. Valenti, B. Spagnolo, G. Basilone, S. Mazzola, S. W. Zgoz, et al., Dynamics of two picophytoplankton groups in mediterranean sea: Analysis of the deep chlorophyll maximum by a stochastic advection-reaction-diffusion model, PLoS One, 8 (2013), e66765. |
| [7] | G. Denaro, D. Valenti, A. La Cognata, B. Spagnolo, A. Bonanno, G. Basilone, et al., Spatio-temporal behaviour of the deep chlorophyll maximum in Mediterranean Sea: Development of a stochastic model for picophytoplankton dynamics, Ecol. Complex, 13 (2013), 21-34. |
| [8] | D. Valenti, G. Denaro, A. L. Cognata, B. Spagnolo, Picophytoplankton dynamics in noisy marine environment, Acta. Phys. Pol B, 43 (2012), 1227-1240. |
| [9] | A. S. Heiskanen, T. Tamminen, K. Gundersen, Impact of planktonic food web structure on nutrient retention and loss from a late summer pelagic system in the coastal northern Baltic Sea, Mar. Ecol. Prog. Ser., 145 (1996), 195-208. |
| [10] | M. Scheffe, Fish and nutrients interplay determines algal biomass: a minimal model, Oikos, 62 (1991), 271-282. |
| [11] | A. B. Medvinsky, S. V. Petrovskii, I. A. Tikhonova, H. Malchow, B. L. Li, Spatio-temporal complexity of plankton and fish dynamics, SIAM Rev., 44 (2002), 311-370. |
| [12] | P. Panja, S. K. Mondal, Stability analysis of coexistence of three species prey-predator model, Nonlinear Anal., 81 (2015), 373-382. |
| [13] | A. Sharma, A. K. Sharma, K. Agnihotri, Complex dynamic of plankton-fish interaction with quadratic harvesting and time delay, Model Earth Syst. Environ., 2 (2016), 1-17. |
| [14] | X. Y. Meng, Y.Q. Wu, Bifurcation and control in a singular phytoplankton-zooplankton-fish model with nonlinear fish harvesting and taxation, Int. J. Bifurcat. Chaos, 28 (2018), 1850042. |
| [15] | Z. Wei, J. Sugie, Global asymptotic stability and equiasymptotic stability for a time-varying phytoplankton-zooplankton-fish system, Nonlinear Anal. Real World Appl., 46 (2019), 116-136. |
| [16] | B. Buonomo, M. Cerasuolo, The effect of time delay in plant-pathogen interactions with host demography, Math. Biosci. Eng., 12 (2015), 473-490. |
| [17] | B. Tian, Y. Qiu, N. Chen, Periodic and almost periodic solution for a non-autonomous epidemic predator-prey system with time-delay, Appl. Math. Copmut., 215 (2009), 779-790. |
| [18] | O. A. Chichigina, A. A. Dubkov, D, Valenti, B. Spagnolo, Stability in a system subject to noise with regulated periodicity, Phys. Rev. E, 84 (2011), 021134. |
| [19] | D. Valenti, L. Tranchina, M. Brai, A. Caruso, C. Cosentino, B. Spagnolo, Environmental metal pollution considered as noise: Effects on the spatial distribution of benthic foraminifera in two coastal marine areas of Sicily (Southern Italy), Ecol. Model, 213 (2008), 449-462. |
| [20] | A. A. Dubkov, B. Spagnolo, Verhulst model with Lévy white noise excitation, Eur. Phys. J. B, 65 (2008), 361-367. |
| [21] | Q. Luo, X. Mao, Stochastic population dynamics under regime switching, J. Math. Anal. Appl., 355 (2009), 577-593. |
| [22] | M. Slatkin, The dynamics of a Population in a Markovian environment, Ecology, 59 (1978), 249-256. |
| [23] | H. Qiu, W. Deng, Stationary distribution and global asymptotic stability of a three-species stochastic food-chain system, Turk. J. Math., 41 (2017), 1292-1307. |
| [24] | Y. Ma, Q. Zhang, L. Wang, T. Kang, Dissipative control of a three-species food chain stochastic system with a hidden Markovchain, Adv. Differ. Equ-Ny., 2017 (2017), 1-22. |
| [25] | Y. Lin, D. Jiang, Long-time behavior of a stochastic predator-prey model with modified Leslie-Gower and Holling-type II schemes, Int. J. Biomath, 9 (2016), 1650039. |
| [26] | J. Lv, K. Wang, Asymptotic properties of a stochastic predator-prey system with Holling II functional response, Commun. Nonlinear. Sci. Numer Simulat., 16 (2011), 4037-4048. |
| [27] | Z. Liu, N. Shi, D. Jiang, C. Ji, The Asymptotic behavior of a stochastic predator-prey system with Holling II functional response, Abstr. Appl. Anal., 2012 (2012), 1-14. |
| [28] | G. Gilioli, S. Pasquali, F. Ruggeri, Nonlinear functional response parameter estimation in a stochastic predator-prey model, Math. Biosci. Eng., 9 (2012), 75-96. |
| [29] | C. S. Reynolds, The Ecology of Freshwater Phytoplankton, Cambridge University Press, Cambridge, 1984. |
| [30] | C. W. Clark, Mathematical Bio-Economics: The Optimal Management of Renewable Resources, Wiley, New York, 1976. |
| [31] | C. W. Clark, Bioeconomic Modeling and Resource Management, in Applied Mathematical Ecology (eds. S. A. Levin), Springer, 1989, 11-57. |
| [32] | M. Mesterton-Gibbons, On the optimal policy for combining harvesting of predator and prey, Nat. Resour. Model, 3 (1988), 63-90. |
| [33] | M. Mesterton-Gibbons, A technique for finding optimal two-species harvesting policies, Ecol. Model, 92 (1996), 235-244 |
| [34] | S. Wang, L. Wang, T. Wei, Optimal harvesting for a stochastic predator-prey model with S-type distributed time delays, Methodol Comput. Appl., 20 (2016), 37-68. |
| [35] | M. Liu, C. Bai, Analysis of a stochastic tri-trophic food-chain model with harvesting, J. Math. Biol., 73 (2016), 597-625. |
| [36] | M. Liu, X. He, J. Yu, Dynamics of a stochastic regime-switching predator-prey model with harvesting and distributed delay, Nonlinear Anal. Hybrid Syst., 28 (2018), 87-104. |
| [37] | W. J. Anderson, Continuous-Time Markov Chains, Springer, New York, 1991. |
| [38] | R. M. May, Stability and Complexity in Model Ecosystems, Princeton University Press, Princeton, 1975. |
| [39] | X. Zhang, W. Li, M. Liu, K. Wang, Dynamics of a stochastic Holling II one-predator two-prey system with jumps, Phys. A, 421 (2015), 571-582. |
| [40] | X. Mao, C. Yuan, Stochastic Differential Equations with Markovian Switching, Imperial College Press, London, 2006. |
| [41] | M. Liu, J. Yu, P. S. Mandal, Dynamics of a stochastic delay competitive model with harvesting and Markovian switching, Appl. Math. Comput., 337 (2018), 335-349. |
| [42] | V. M. Popov, Hyperstability of Control Systems, Springer-Verlag, New York, 1973. |
| [43] | D. Prato, J. Zabczyk, Ergodicity for Infinite Dimensional Systems, Cambridge University Press, Cambridge, 1996. |
| [44] | L. Thomas, Estimating Phytoplankton Growth Rates from Compositional Data, in Oceanography/Biological Oceanography Massachusetts Institute of Technology and Woods Hole Oceanographic Institution, Massachusetts Institute Of Technology, 2008. |
| [45] | T. Nanazato, M. Yasuno, Population dynamics and production of cladoceran zooplankton in the highly eutrophic Lake Kasumigaura, Hydrobiologia, 124 (1981), 13-22. |
| [46] | Y. Wang, Q. Liu, Estimating natural mortality from stock size and catch data (in Chinese), Period. Ocean Univ. China, 35 (2005), 020-024. |
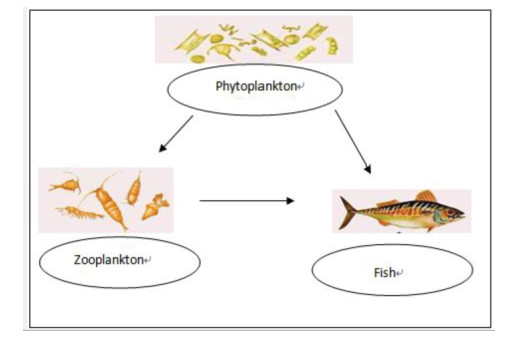
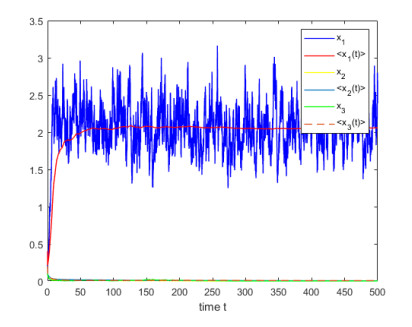
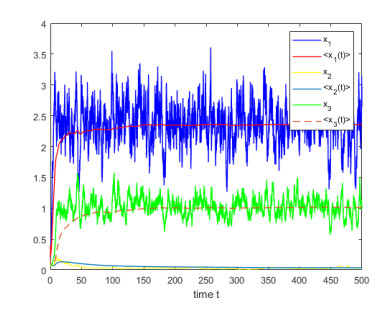
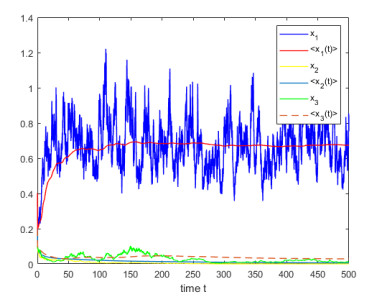
Figures(4)

Yuanpei Xia, Weisong Zhou, Zhichun Yang. Global analysis and optimal harvesting for a hybrid stochastic phytoplankton-zooplankton-fish model with distributed delays[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 6149-6180. doi: 10.3934/mbe.2020326







 DownLoad:
DownLoad: