Citation: Andrés Donoso-Bravo, Pedro Gajardo, Matthieu Sebbah, Diego Vicencio. Comparison of performance in an anaerobic digestion process: one-reactor vs two-reactor configurations[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2447-2465. doi: 10.3934/mbe.2019122

| [1] | M. Cooney, N. Maynard and C. Cannizzaro, et al., Two-phase anaerobic digestion for production of hydrogen-methane mixtures, Biores. Technol., 98 (2007), 2641–2651. |
| [2] | R. Li, S. Chen and X. Li, Biogas production from anaerobic co-digestion of food waste with dairy manure in a two-phase digestion system, App. Biochem. Biotechnol., 160 (2010), 643–654. |
| [3] | D. J. Batstone, J. Keller and I. Angelidaki, et al., The iwa anaerobic digestion model no 1 (adm1), Water Sci. Technol., 45 (2002), 65–73. |
| [4] | O. Bernard, Z. Hadj-Sadok and D. Dochain, et al., Dynamical model development and parameter identification for an anaerobic wastewater treatment process, Biotechnol. Bioeng., 75 (2001), 424– 438. |
| [5] | M. Weedermann, G. S. Wolkowicz and J. Sasara, Optimal biogas production in a model for anaerobic digestion, Nonlinear Dynam., 81 (2015), 1097–1112. |
| [6] | Y. Daoud, N. N. Abdellatif and J. Harmand, Modèles mathématiques de digestion anaérobie: effet de l'hydrolyse sur la production du biogaz, 2017. https://hal.archives-ouvertes. fr/hal-01562353, Working paper or preprint. |
| [7] | M. Sbarciog, M. Loccufier and A. Vande Wouwer, On the optimization of biogas production in anaerobic digestion systems, IFAC Proceed. Vol., 44 (2011), 7150–7155. |
| [8] | M. Sbarciog, M. Loccufier and A. Vande Wouwer, An optimizing start-up strategy for a biomethanator, Bioprocess Biosys. Eng., 35 (2012), 565–578. |
| [9] | M. Sbarciog, J. A. Moreno and A. Vande Wouwer, A biogas-based switching control policy for anaerobic digestion systems1, IFAC Proceed. Vol., 45 (2012), 603–608. |
| [10] | B. Benyahia, T. Sari and B. Cherki, et al., Bifurcation and stability analysis of a two step model for monitoring anaerobic digestion processes, J. Process Control, 22 (2012), 1008–1019. |
| [11] | H. L. Smith and P. Waltman, The theory of the chemostat, vol. 13 of Cambridge Studies in Mathematical Biology, Cambridge University Press, Cambridge, 1995, Dynamics of microbial competition. |
| [12] | T. Bayen and P. Gajardo, On the steady state optimization of the biogas production in a two-stage anaerobic digestion model, J. Mathemat. Biol., 78 (2019), 1067–1087. |
| [13] | T. Bayen, O. Cots and P. Gajardo, Analysis of an Optimal Control Problem Related to the Anaerobic Digestion Process, J. Optim. Theory Appl., 178 (2018), 627–659. |
| [14] | K. Postawa, Novel solutions in modeling of anaerobic digestion process-two-phase AD models development and comparison, In. J. Chemical React. Eng., 16. |
| [15] | Z. Chen, D. Hu and Z. Zhang, et al., Modeling of two-phase anaerobic process treating traditional chinese medicine wastewater with the IWA Anaerobic Digestion Model No. 1, Biores. Technol., 100 (2009), 4623–4631. http://www.sciencedirect.com/science/article/pii/ S0960852409004830. |
| [16] | A. Novick and L. Szilard, Experiments with the chemostat on spontaneous mutations of bacteria, Proceed. Nat. Aca. Sci., 36 (1950), 708–719. |
| [17] | J. Heßeler, J. K. Schmidt and U. Reichl, et al., Coexistence in the chemostat as a result of metabolic by-products, J. Math. Biol., 53 (2006), 556–584. |
| [18] | J. Harmand, C. Lobry and A. Rapaport, et al., Le chémostat: Théorie mathématique de la culture continue de micro-organismes, vol. 1, ISTE Editions, 2017. |
| [19] | L. Baere and B. Mattheeuws, Anaerobic digestion of the organic fraction of municipal solid waste in Europe: Status, experience and prospects, Waste Management, 3 (2012), 517–526. |
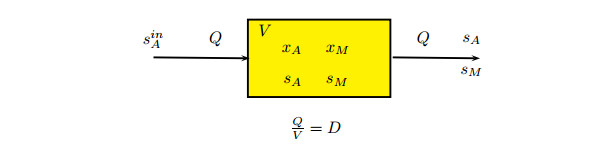
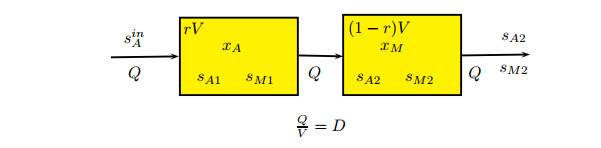
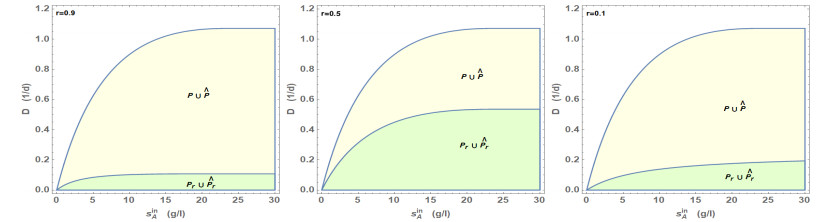
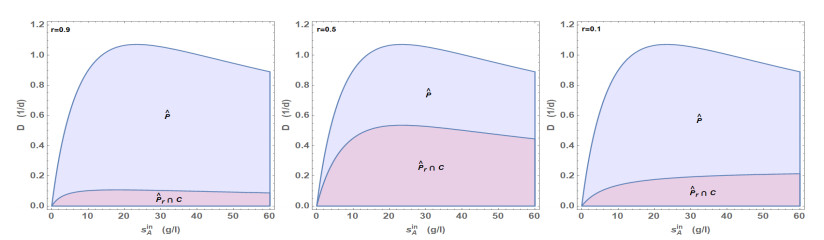
Figures(5) / Tables(1)

Andrés Donoso-Bravo, Pedro Gajardo, Matthieu Sebbah, Diego Vicencio. Comparison of performance in an anaerobic digestion process: one-reactor vs two-reactor configurations[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2447-2465. doi: 10.3934/mbe.2019122







 DownLoad:
DownLoad: