Citation: Yanqin Wang, Xin Ni, Jie Yan, Ling Yang. Modeling transcriptional co-regulation of mammalian circadian clock[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1447-1462. doi: 10.3934/mbe.2017075

| [1] | [ M. Adimy,F. Crauste,S. G. Ruan, Periodic oscillations in leukopoiesis models with two delays, Journal of Theoretical Biology, 242 (2006): 288-299. |
| [2] | [ M. P. Antoch,V. Y. Gorbacheva,O. Vykhovanets,A. Y. Nikitin, Disruption of the circadian clock due to the Clock mutation has discrete effects on aging and carcinogenesis, Cell Cycle, 7 (2008): 1197-1204. |
| [3] | [ D. B. Forger,C. S. Peskin, A detailed predictive model of the mammalian circadian clock, Proceedings of the National Academy of Sciences of the United States of America, 100 (2003): 14806-14811. |
| [4] | [ A. Goldbeter, A model for circadian oscillations in the Drosophila period protein (PER), Proceedings. Biological sciences / The Royal Society, 261 (1995): 319-324. |
| [5] | [ A. Goldbeter, Biochemical Oscillations and Cellular Rhythms: The Molecular Bases of Periodic and Chaotic Behaviour, Cambridge University Press, 1996. |
| [6] | [ C. I. Hong,J. J. Tyson, A proposal for temperature compensation of the circadian rhythm in Drosophila based on dimerization of the per protein, Chronobiology International, 14 (1997): 521-529. |
| [7] | [ J. K. Kim and D. B. Forger, A mechanism for robust circadian timekeeping via stoichiometric balance, Molecular Systems Biology, 8 (2012), 630. |
| [8] | [ R. V. Kondratov,A. A. Kondratova,V. Y. Gorbacheva, Early aging and age-related pathologies in mice deficient in BMAL1, the core component of the circadian clock, Genes & Developoment, 20 (2006): 1868-1873. |
| [9] | [ C. C. Lee, Tumor suppression by the mammalian Period genes, Cancer Causes Control, 17 (2006): 525-530 [PubMed: 16596306]. |
| [10] | [ J. C. Leloup,A. Goldbeter, Toward a detailed computational model for the mammalian circadian clock: Sensitivity analysis and multiplicity of oscillatory mechanisms, J. Theoret. Biol., 230 (2004): 541-562. |
| [11] | [ P. L. Lowrey,J. S. Takahashi, Mammalian circadian biology: elucidating genome-wide levels of temporal organization, Annual Review of Genomics and Human Genetics, 5 (2004): 407-441. |
| [12] | [ H. P. Mirsky,A. C. Liu,D. K. Welsh,S. A. Kay,F. J. Doyle, A model of the cell-autonomous mammalian circadian clock, Proceedings of the National Academy of Sciences of the United States of America, 106 (2009): 11107-11112. |
| [13] | [ S. G. Ruan,J. J. Wei, On the zeros of transcendental functions with applications to stability of delay differential equations with two delays, Dynamics of Continuous, Discrete and Impulsive Systems, Series A: Mathematical Analysis, 10 (2003): 863-874. |
| [14] | [ F. A. Scheer,M. F. Hilton,C. S. Mantzoros,S. A. Shea, Adverse metabolic and cardiovascular consequences of circadian misalignment, Proceedings of the National Academy of Sciences of the United States of America, 106 (2009): 4453-4458. |
| [15] | [ J. J. Tyson,C. I. Hong,C. D. Thron,B. Novak, A simple model of circadian rhythms based on dimerization and proteolysis of PER and TIM, Biophys Journal, 77 (1999): 2411-2417. |
| [16] | [ M. Ukai-Tadenuma,R. G. Yamada,H. Xu,J. A. Ripperger,A. C. Liu,H. R. Ueda, Delay in feedback repression by cryptochrome 1 is required for circadian clock function, Cell, 144 (2011): 268-281. |
| [17] | [ J. Yan,G. Shi,Z. Zhang,X. Wu,Z. Liu,L. Xing,Z. Qu,Z. Dong,L. Yang,Y. Xu, An intensity ratio of interlocking loops determines circadian period length, Nucleic Acids Research, 42 (2014): 10278-10287. |
| [18] | [ X. Yang,M. Downes,R. T. Yu,A. L. Bookout,W. He,M. Straume,D. J. Mangelsdorf,R. M. Evans, Nuclear receptor expression links the circadian clock to metabolism, Cell, 126 (2006): 801-810. |
| [19] | [ W. Yu,M. Nomura,M. Ikeda, Interactivating feedback loops within the mammalian clock: BMAL1 is negatively autoregulated and upregulated by CRY1, CRY2, and PER2, Biochemical and Biophysical Research Communications, 290 (2002): 933-941. |
| [20] | [ E. E. Zhang,S. A. Kay, Clocks not winding down: Unravelling circadian networks, Nature Reviews Molecular Cell Biology, 11 (2010): 764-776. |
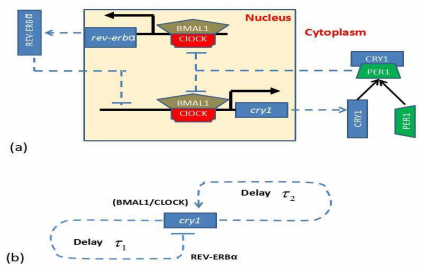
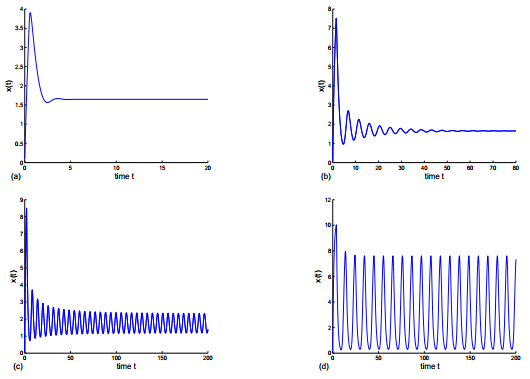
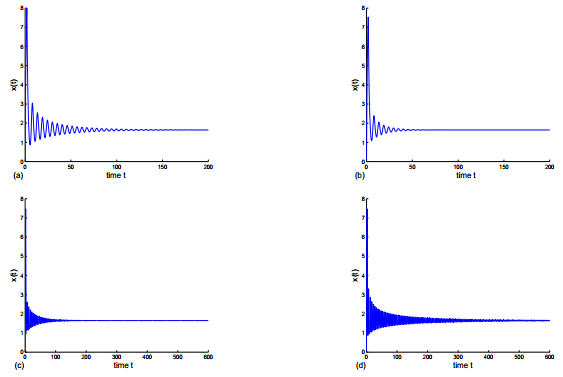
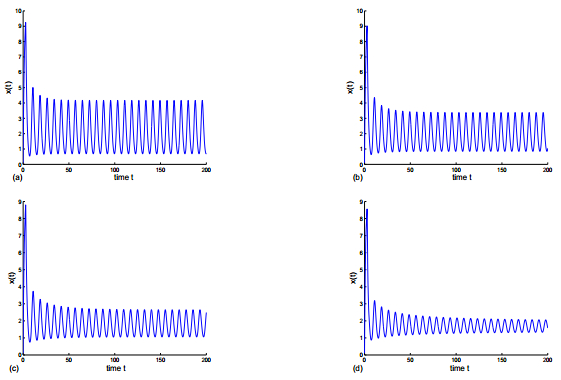
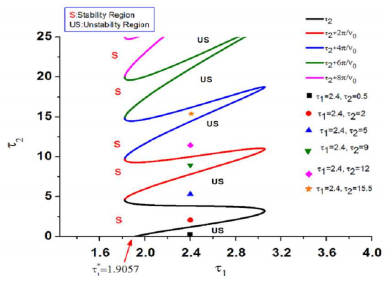
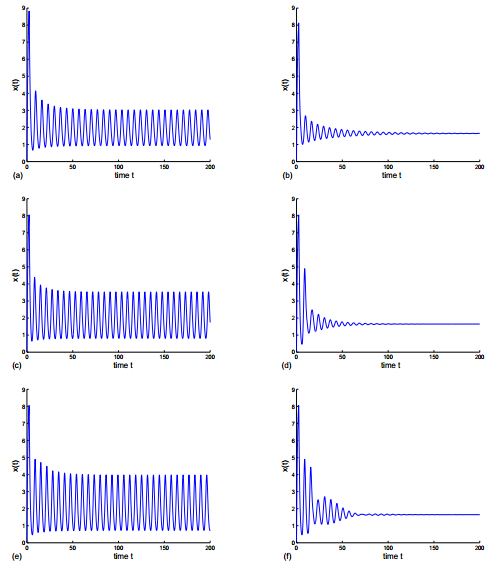
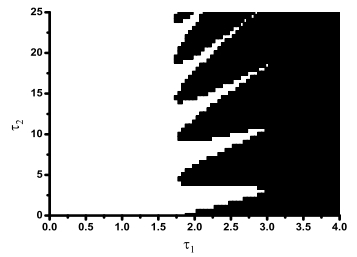
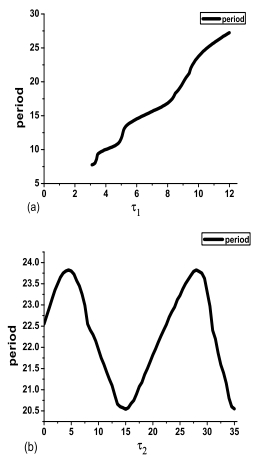
Figures(8)

Yanqin Wang, Xin Ni, Jie Yan, Ling Yang. Modeling transcriptional co-regulation of mammalian circadian clock[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1447-1462. doi: 10.3934/mbe.2017075







 DownLoad:
DownLoad: