Citation: Binaya Tuladhar, Hana M. Dobrovolny. Testing the limits of cardiac electrophysiology models through systematic variation of current[J]. AIMS Mathematics, 2020, 5(1): 140-157. doi: 10.3934/math.2020009

| [1] | E. Anyukhovsky and M. Rosen, Electrophysiologic effects of alprafenone on canine cardiac tissue, J. Cardiovasc. Pharm., 24 (1994), 411-419. |
| [2] | R. Atanasiu, L. Gouin, M. A. Mateescu, et al. Class Ⅲ antiarrhythmic effects of ceruloplasmin on rat heart, Can. J. Physiol. Pharm., 74 (1999), 652-656. |
| [3] | T. Bányász, L. Bárándi, G. Harmati, et al. Mechanism of reverse rate-dependent action of cardioactive agents, Curr. Med. Chem., 18 (2011), 3597-3606. |
| [4] | T. Banyasz, B. Horvath, L. Virag, et al. Reverse rate dependency is an intrinsic property of canine cardiac preparations, Cardiovasc. Res., 84 (2009), 237-244. |
| [5] | L. Bárándi, L. Virág, N. Jost, et al. Reverse rate-dependent changes are determined by baseline action potential duration in mammalian and human ventricular preparations, Basic Res. Cardiol., 105 (2010), 315-323. |
| [6] | G. Beatch, D. Davis, S. Laganiere, et al. Rate-dependent effects of sematilide on ventricular monophasic action potential duration and delayed rectifier K+ current in rabbits, J. Cardiovasc. Pharm., 28 (1996), 618-630. |
| [7] | O. Bernus, R. Wilder, C. W. Zemlin, et al. A computationally efficient electrophysiological model of human ventricular cells, American Journal of Physiology-Heart and Circulatory Physiology, 282 (2002), 2296-2308. |
| [8] | K. Blinova, Q. Dang, D. Millard, et al. International multisite study of human-induced pluripotent stem cell-derived cardiomyocytes for drug proarrhythmic potential assessment, Cell Rep., 24 (2018), 3582-3592. |
| [9] | O. J. Britton, A. Bueno-Orovio, K. V. Ammel, et al. Experimentally calibrated population of models predicts and explains intersubject variability in cardiac cellular electrophysiology, Proceedings of the National Academy of Sciences of the United States of America, 110 (2013), E2098-E2105. |
| [10] | D. I. Cairns, F. H. Fenton and E. M. Cherry, Efficient parameterization of cardiac action potential models using a genetic algorithm, Chaos, 27 (2017), 093922. |
| [11] | T. J. Campbell, K. R. Wyse and R. Pallandi, Differential effects on action potential duration of class Ia, b and c antiarrhythmic drugs: modulation by stimulation rate and extracellular K+ concentration, Clin. Exp. Pharmacol. P., 18 (1991), 533-541. |
| [12] | J. Carro, J. F. Rodriguez-Matas, V. Monasterio, et al. Limitations in electrophysiological model development and validation caused by differences between simulations and experimental protocols, Prog. Biophys. Mol. Bio., 129 (2017), 53-64. |
| [13] | A. Carusi, K. Burrage and B. Rodriguez, Bridging experiments, models and simulations: an integrative approach to validation in computational cardiac electrophysiology, Am. J. Physiol.-Heart C., 303 (2012), H144-H155. |
| [14] | I. Cavero and H. Holzgrefe, Comprehensive in vitro proarrhythmia assay, a novel in vitro/in silico paradigm to detect ventricular proarrhythmic liability: a visionary 21st century initiative, Expert Opin. Drug Saf., 13 (2014), 745-758. |
| [15] | C.-E. Chiang, H.-N. Luk, T.-M. Wang, et al. Effects of sildenafil on cardiac repolarization, Cardiovasc. Res., 255 (2002), 290-299. |
| [16] | R. Clayton, O. Bernus, E. Cherry, et al. Models of cardiac tissue electrophysiology: Progress, challenges and open questions, Prog. Biophys. Mol. Bio., 104 (2011), 22-48. |
| [17] | J. Cooper, A. Corrias, D. Gavaghan, et al. Considerations for the use of cellular electrophysiology models within cardiac tissue simulations, Prog. Biophys. Mol. Bio., 107 (2011), 74-80. |
| [18] | D.-Z. Dai, F. Yu, H.-T. Li, et al. Blockade on sodium, potassium, and calcium channels by a new antiarrhythmic agent CPU 86017, Drug Develop. Res., 39 (1996), 138-146. |
| [19] | A. C. Daly, M. Clerx, K. A. Beattie, et al. Reproducible model development in the cardiac electrophysiology web lab, Prog. Biophys. Mol. Bio., 139 (2018), 3-14. |
| [20] | R. A. Devenyi, F. A. Ortega, W. Groenendaal, et al. Differential roles of two delayed rectifier potassium currents in regulation of ventricular action potential duration and arrhythmia susceptibility, J. Physiol., 595 (2017), 2301-2317. |
| [21] | H. J. Duff, R. S. Sheldon and N. J. Cannon, Tetrodotoxin: Sodium channel specific antiarrhythmic activity, Cardiovasc. Res., 22 (1988), 800-807. |
| [22] | J. Eastman, J. Sass, J. M. Gomes, et al. Using delay differential equations to induce alternans in a model of cardiac electrophysiology, J. Theor. Biol., 404 (2016), 262-272. |
| [23] | B. Fermini, N. Jurkiewicz, B. Jow, et al. Use-dependent effects of the class-Ⅲ antiarrhythmic agent NE-10064 (azimilide) on cardiac repolarization — block of delayed rectifier potassium and L-type calcium currents, J. Cardiovasc. Pharm., 26 (1995), 259-271. |
| [24] | J. J. Fox, J. L. McHarg and R. F. Gilmour, Ionic mechanism of electrical alternans, Am. J. Physiol.-Heart C., 282 (2002), H516-H530. |
| [25] | K. Fukuda, J. Watanabe, T. Yagi, et al. A sodium channel blocker, pilsicainide, produces atrial post-repolarization refractoriness through the reduction of sodium channel availability, Tohoku J. Exp. Med., 225 (2011), 35-42. |
| [26] | L. Geng, C.-W. Kong, A. O. Wong, et al. Probing flecainide block of I-Na using human pluripotent stem cell-derived ventricular cardiomyocytes adapted to automated patch-clamping and 2D monolayers, Toxicol. Lett., 294 (2018), 61-72. |
| [27] | J. K. Gibson, Y. Yue, J. Bronson, et al. Human stem cell-derived cardiomyocytes detect drugmediated changes in action potentials and ion currents, J. Pharmacol. Tox. Met., 70 (2014), 255-267. |
| [28] | G. Gintant, The class Ⅲ effect of azimilide is not associated with reverse use-dependence in open-chest dogs, J. Cardiovasc. Pharm., 31 (1998), 945-953. |
| [29] | R. A. Gray and P. Pathmanathan, Patient-specific cardiovascular computational modeling: Diversity of personalization and challenges, J. Cardiovasc. Trans. Res., 11 (2018), 80-88. |
| [30] | W. Groenendaal, F. A. Ortega, A. R. Kherlopian, et al. Cell-specific cardiac electrophysiology models, PLoS Comput. Biol., 11 (2015), e1004242. |
| [31] | R. N. Gutenkunst, J. J. Waterfall, F. P. Casey, et al. Universally sloppy parameter sensitivities in systems biology models, PLoS Comput. Biol., 3 (2007), e189. |
| [32] | G. Hall, S. Bahar and D. Gauthier, Prevalence of rate-dependent behaviors in cardiac muscle, Phys. Rev. Lett., 82 (1999), 2995-2998. |
| [33] | J. Heijman, S. Ghezelbash and D. Dobrev, Investigational antiarrhythmic agents: promising drugs in early clinical development, Expert Opin. Inv. Drug., 26 (2017), 897-907. |
| [34] | A. Hodgkin and A. Huxley, A quantitative description of membrane current and its application to conduction and excitation in nerve, J. Physiol., 117 (1952), 500-544. |
| [35] | J. W. Holmes and J. Lumens, Clinical applications of patient-specific models: The case for a simple approach, J. Cardiovasc. Trans. Res., 11 (2010), 71-79. |
| [36] | X. Huang, Y. Qian, X. Zhang, et al. Hysteresis and bistability in periodically paced cardiac tissue, Phys. Rev. E, 81 (2010), 051903. |
| [37] | I. Jacobson, G. Duker, M. Florentzson, et al. Experimental study electrophysiological characterization and antiarrhythmic efficacy of the mixed potassium channel-blocking antiarrhythmic agent AZ13395438 in vitro and in vivo, J. Cardiovasc. Pharmacol. Ther., 1 (2013), 290-300. |
| [38] | D. Jans, G. C. ad Olga Krylychkina, L. Hoffman, et al. Action potential-based MEA platform for in vitro screening of drug-induced cardiotoxicity using human iPSCs and rat neonatal myocytes, J. Pharmacol. Toxicol. Meth., 1 (2017), 290-300. |
| [39] | Q. Jin, X. Chen, W. M. Smith, et al. Effects of procainamide and sotalol on restitution properties, dispersion of refractoriness, and ventricular fibrillation activation patterns in pigs, J. Cardiovasc. Pharm., 19 (2008), 1090-1097. |
| [40] | Q. Jin, J. Zhou, N. Zhang, et al. Ibutilide decreases defibrillation threshold by the reduction of activation pattern complexity during ventricular fibrillation in canine hearts, Chinese Med. J., 125 (2012), 2701-2707. |
| [41] | N. K. Jurkiewicz and M. C. Sanguinetti, Rate-dependent prolongation of cardiac action potentials by a methanesulfonanilide class Ⅲ antiarrhythmic agent. Specific block of rapidly activating delayed rectifier K+ current by dofetilide, Circ. Res., 72 (1993), 75-83. |
| [42] | S. Kalb, H. Dobrovolny, E. Tolkacheva, et al. The restitution portrait: A new method of investigating rate-dependent restitution, J. Cardiovasc. Electr., 15 (2004), 698-709. |
| [43] | S. Kalb, J. Fox, E. Tolkacheva, et al. Parameter estimation in mapping models with memory, Anatomical Record A, 288 (2006), 579-86. |
| [44] | R. E. Klabunde, Cardiovascular Physiological Concepts, Lippincott Williams & Wilkins, Philadelphia, 2011. |
| [45] | I. Kodama, K. Kamiya and J. Toyama, Cellular electropharmacology of amiodarone, Cardiovasc. Res., 35 (1997), 13-29. |
| [46] | J.-Y. Le Guennec, J. Thireau, A. Ouille, et al. Inter-individual variability and modeling of electrical activity: a possible new approach to explore cardiac safety? Sci. Rep., 6 (2016), 37948. |
| [47] | L. Livshitz and Y. Rudy, Uniqueness and stability of action potential models during rest, pacing, and conduction using problem-solving environment, Biophys. J., 97 (2009), 1265-1276. |
| [48] | H. R. Lu, E. Vlaminckx, A. Teisman, et al. Choice of cardiac tissue plays an important role in the evaluation of drug-induced prolongation of the QT interval in vitro in rabbit, J. Pharmacol. Tox. Met., 52 (2005), 90-105. |
| [49] | D. lu Bai, W. zhou Chen, Y. xin Bo, et al. Discovery of N-(3, 5-bis(1-pyrrolidylmethyl)-4-hydroxy-benzyl)-4-methoxybenzenesulfamide (sulcardine) as a novel anti-arrhythmic agent, Acta Pharmacol. Sin., 33 (2012), 1176-1186. |
| [50] | H. Marschang, T. Beyer, L. Karolyi, et al. Differential rate and potassium dependent effects of class Ⅲ agents d-sotalol and dofetilide on guinea pig papillary muscle, Cardiovasc. Drug. Ther., 12 (1998), 573-583. |
| [51] | G. R. Mirams, M. R. Davies, Y. Cui, et al. Application of cardiac electrophysiology simulations to proarrhythmic safety testing, Brit. J. Pharmacol., 167 (2012), 932-945. |
| [52] | A. Muszkiewicz, O. J. Britton, P. Gemmell, et al. Variability in cardiac electrophysiology: Using experimentally-calibrated populations of models to move beyond the single virtual physiological human paradigm, Prog. Biophys. Mol. Biol., 120 (2016), 932-945. |
| [53] | S. Narayan, T-wave alternans and the susceptibility to ventricular arrhythmias, J. Am. Coll. Cardiol., 47 (2006), 269-281. |
| [54] | H. B. Ni, S. Morotti and E. Grandi, A heart for diversity: Simulating variability in cardiac arrhythmia research, Front. Physiol., 9 (2018), 958. |
| [55] | D. Noble, Modification of Hodgkin-Huxley equations applicable to Purkinje fibre action and pace-maker potentials, J. Physiol., 160 (1962), 317-52. |
| [56] | D. Noble, A. Garny and P. J. Noble, How the Hodgkin-Huxley equations inspired the Cardiac Physiome Project, J. Physiol., 590 (2012), 2613-2628. |
| [57] | K. Noguchi, J. Kase, M. Saitoh, et al. Effects of HNS-32, a novel antiarrhythmic agent, on guineapig myocardium, Pharmacology, 64 (2002), 36-42. |
| [58] | R. A. Oliver, G. M. Hall, S. Bahar, et al. Existence of bistability and correlation with arrhythmogenesis in paced sheep atria, J. Cardiovasc. Electr., 11 (2000), 797-805. |
| [59] | R. A. Oliver, C. S. Henriquez and W. Krassowska, Bistability and correlation with arrhythmogenesis in a model of the right atrium, Ann. Biomed. Eng., 33 (2005), 577-589. |
| [60] | C. Omichi, S. Zhou, M. Lee, et al. Effects of amiodarone on wave front dynamics during ventricular fibrillation in isolated swine right ventricle, Am. J. Physiol.-Heart. C., 282 (2002), H1063-H1070. |
| [61] | P. M. Orth, J. C. Hesketh, C. K. Mak, et al. RSD1235 blocks late INa and suppresses early afterdepolarizations and torsades de pointes induced by class Ⅲ agents, Cardiovasc. Res., 70 (2006), 486-496. |
| [62] | O. E. Osadchii, Dofetilide promotes repolarization abnormalities in perfused guinea-pig heart, Cardiovasc. Drug. Ther., 26 (2012), 489-500. |
| [63] | O. E. Osadchii, Quinidine elicits proarrhythmic changes in ventricular repolarization and refractoriness in guinea-pig, Can. J. Physiol. Pharmacol., 91 (2013), 306-315. |
| [64] | O. E. Osadchii, Effects of antiarrhythmics on the electrical restitution in perfused guinea-pig heart are critically determined by the applied cardiac pacing protocol, Exp. Physiol., 104 (2019), 490-504. |
| [65] | T. Osaka, E. Yokoyama, H. Hasebe, et al. Effects of chronic amiodarone on the electrical restitution in the human ventricle with reference to its antiarrhythmic efficacy, J. Cardiovasc. Eletr., 22 (2011), 669-676. |
| [66] | T. Osaka, E. Yokoyama, Y. Kushiyama, et al. Opposing effects of bepridil on ventricular repolarization in humans-inhomogeneous prolongation of the action potential duration vs flattening of its restitution kinetics, Circ. J., 73 (2009), 1612-1618. |
| [67] | M. Pan, P. J. Gawthrop, K. Tran, et al. Bond graph modelling of the cardiac action potential: implications for drift and non-unique steady states, Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 474 (2018), 20180106. |
| [68] | C. Pankusci, T. Banyasz, J. Magyar, et al. Electrophysiological effects of EGIS-7229, a new antiarrhythmic agent, in isolated mammalian and human cardiac tissues, N-S Arch. Pharmacol., 355 (1997), 398-405. |
| [69] | J. M. Pastore, S. D. Girouard, K. R. Laurita, et al. Mechanism linking T-wave alternans to the genesis of cardiac fibrillation, Circulation, 99 (1999), 1385-1394. |
| [70] | P. Pathmanathan and R. A. Gray, Validation and trustworthiness of multiscale models of cardiac electrophysiology, Front. Physiol., 9 (2018), 106. |
| [71] | A. Pezhouman, S. Madahian, H. Stepanyan, et al. Selective inhibition of late sodium current suppresses ventricular tachycardia and fibrillation in intact rat hearts, Heart Rhythm, 11 (2014), 492-501. |
| [72] | A. S. Pickoff and A. Stolfi, Comparison of the rate dependent effects of dofetilide and ibutilide in the newborn heart, Pacing and Clinical Electrophysiology, 24 (2001), 816-823. |
| [73] | S. Polak, B. Wisniowska, K. Fijorek, et al. In vitro-in vivo extrapolation of drug-induced proarrhythmia predictions at the population level, General Pharmacology, 19 (2014), 275-281. |
| [74] | X. Qi, D. Newman and P. Dorian, The class Ⅲ effect of azimilide is not associated with reverse use-dependence in open-chest dogs, J. Cardiovasc. Pharm., 34 (1999), 898-903. |
| [75] | D. M. Roden, Antiarrhythmic drugs: from mechanisms to clinical practice, Heart, 84 (2000), 339-346. |
| [76] | D. S. Rosenbaum, L. E. Jackson, J. M. Smith, et al. Electrical alternans and vulnerability to ventricular arrhythmias, The New England Journal of Medicine, 330 (1994), 235-241. |
| [77] | P. T. Sager and M. Behboodikhah, Frequency-dependent electrophysiologic effects of d, l-sotalol and quinidine and modulation by beta-adrenergic stimulation, J. Cardiovasc. Pharm., 25 (1995), 1006-1011. |
| [78] | C. Sánchez, A. Bueno-Orovio, E. Wettwer, et al. Inter-subject variability in human atrial action potential in sinus rhythm versus chronic atrial fibrillation, PLoS ONE, 9 (2014), e105897. |
| [79] | A. A. Sher, K. Wang, A. Wathen, et al. A local sensitivity analysis method for developing biological models with identifiable parameters: Application to cardiac ionic channel modelling, Future Gener. Comp. Sy., 29 (2013), 591-598. |
| [80] | P. K. Shreenivasaiah, S.-H. Rho, T. Kim, et al. An overview of cardiac systems biology, J. Mol. Cell. Cardiol., 44 (2008), 460-469. |
| [81] | J. M. Smith, S. M. Clancy, R. Valeri, et al. Electrical alternans and cardiac electrical instability, Circulation, 77 (1988), 110-21. |
| [82] | E. A. Sosunov, E. P. Anyukhovsky and M. R. Rosen, Effects of quinidine on repolarization in canine epicardium, midmyocardium, and endocardium, Circulation, 96 (1997), 4011-4018. |
| [83] | M.-J. Su, G.-J. Chang, M.-H. Wu, et al. Electrophysiological basis for the antiarrhythmic action and positive inotropy of HA-7, a furoquinoline alkaloid derivative, in rat heart, Brit. J. Pharmacol., 122 (1997), 1285-1298. |
| [84] | J. Takacs, N. Iost, C. Lengyel, et al. Multiple cellular electrophysiological effects of azimilide in canine cardiac preparations, Eur. J. Pharmacol., 470 (2003), 163-170. |
| [85] | E. Tolkacheva, D. Schaeffer, D. Gauthier, et al. Condition for alternans and stability of the 1:1 response pattern in a 'memory' model of paced cardiac dynamics, Phys. Rev. E, 67 (2003), 031904. |
| [86] | M. R. Vagos, H. Arevalo, B. L. de Oliveira, et al. A computational framework for testing arrhythmia marker sensitivities to model parameters in functionally calibrated populations of atrial cells, Chaos, 27 (2017), 093941. |
| [87] | Z. Wang, B. Fermini and S. Nattel, Mechanism of flecainides rate-dependent actions on actionpotential duration in canine atrial tissue, J. Pharmacol. Exp. Ther., 267 (1993), 575-581. |
| [88] | M. Wilhelms, H. Hettmann, M. M. Maleckar, et al. Benchmarking electrophysiological models of human atrial myocytes, Front. Physiol., 3 (2013), 487. |
| [89] | B. A. Williams, D. R. Dickenson and G. N. Beatch, Kinetics of rate-dependent shortening of action potential duration in guinea-pig ventricle; effects of IK1 and IKr blockade, Brit. J. Pharmacol., 126 (1999), 1426-1436. |
| [90] | B. Wisniowska, A. Mendyk, K. Fijorek, et al. Computer-based prediction of the drug proarrhythmic effect: problems, issues, known and suspected challenges, Europace, 16 (2014), 724-735. |
| [91] | R. Wu and A. Patwardhan, Effects of rapid and slow potassium repolarization currents and calcium dynamics on hysteresis in restitution of action potential duration, J. Electrocardiol., 40 (2007), 188-199. |
| [92] | K. Wyse, V. Ye and T. Campbell, Action-potential prolongation exhibits simple dose-dependence for sotalol, but reverse dose-dependence for quinidine and disopyramide — implications for proarrhythmia due to triggered activity, J. Cardiovasc. Pharm., 21 (1993), 316-322. |
| [93] | X. Yang, T. Yu and H. Kesteloot, Clinical and electrophysiologic studies of R61748 (transcainide) — a new class Ic antiarrhythmic drug, Acta Cardiol., 47 (1992), 43-56. |
| [94] | Z. I. Zhu and C. E. Clancy, Genetic mutations and arrhythmia: simulation from DNA to electrocardiogram, J. Electrocardiol., 40 (2007), S47-S50. |
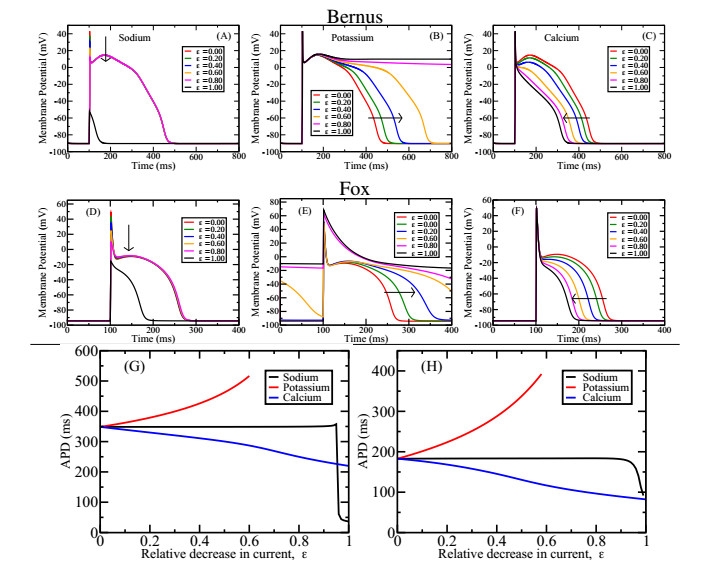
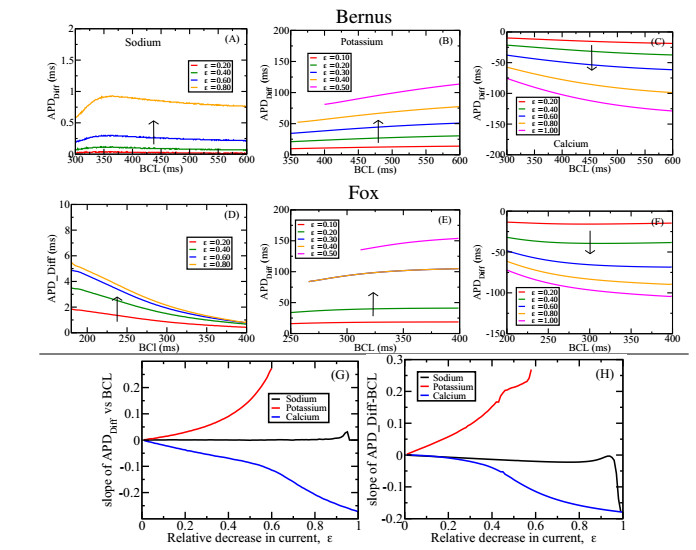
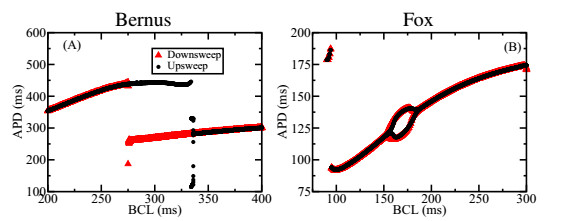
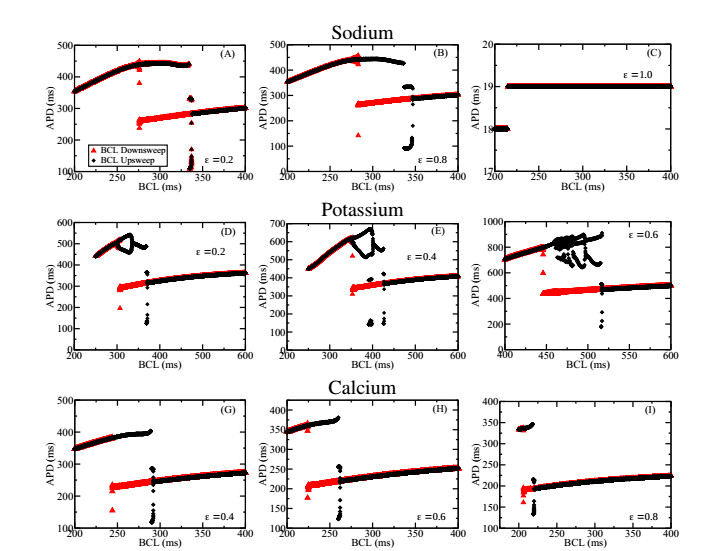
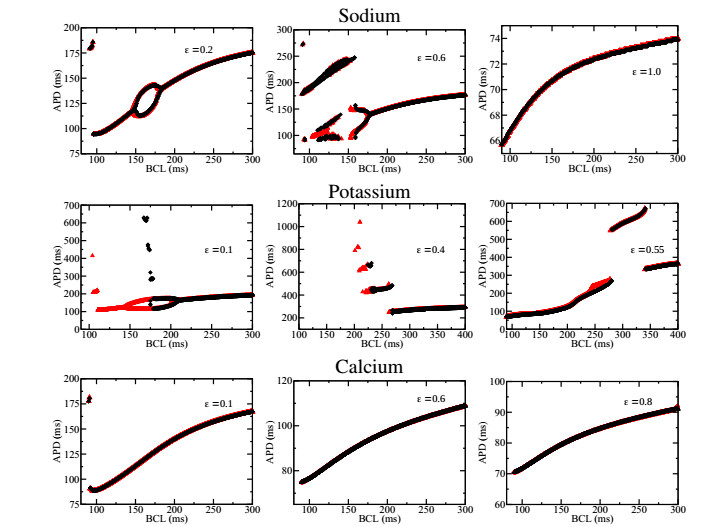
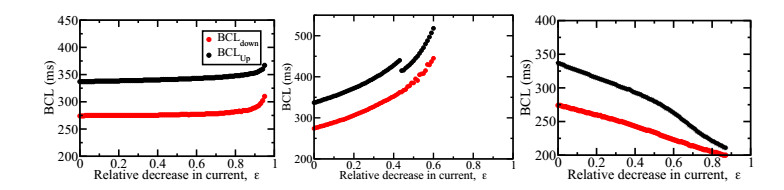
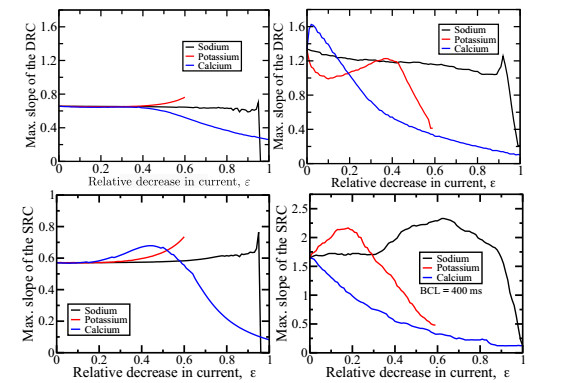
Figures(7)

Binaya Tuladhar, Hana M. Dobrovolny. Testing the limits of cardiac electrophysiology models through systematic variation of current[J]. AIMS Mathematics, 2020, 5(1): 140-157. doi: 10.3934/math.2020009







 DownLoad:
DownLoad: