Citation: Satbir Singh, Peter J. Adams, Albert A. Presto. Simulations of vehicle-induced mixing and near-road aerosol microphysics using computational fluid dynamics[J]. AIMS Environmental Science, 2018, 5(5): 315-339. doi: 10.3934/environsci.2018.5.315

| [1] |
Pope CA, Ezzati M, Dockery DW (2009) Fine-particulate air pollution and life expectancy in the United States. N Engl J Med 360: 376–386. doi: 10.1056/NEJMsa0805646

|
| [2] | Adams PJ, Seinfeld JH (2003) Disproportionate impact of particulate emissions on global cloud condensation nuclei concentrations. Geophys Res Lett 30: 335–343. |
| [3] |
Kanakidou M, Seinfeld JH, Pandis SN, et al. (2005) Organic aerosol and global climate modeling: a review. Atmos Chem Phys 5: 1053–1123. doi: 10.5194/acp-5-1053-2005
|
| [4] |
Spracklen DV, Carslaw KS, Poschl U, et al. (2011) Global cloud condensation nuclei influenced by carbonaceous combustion aerosol. Atmos Chem Phys 11: 9067–9087. doi: 10.5194/acp-11-9067-2011
|
| [5] |
Karner AA, Eisinger DS, Niemeier DA (2010) Near-roadway air quality: synthesizing the findings from real-world data. Environ Sci Technol 44: 5334–5344. doi: 10.1021/es100008x
|
| [6] |
Padro-Martinez LT, Patton AP, Trull JB, et al. (2012) Mobile monitoring of particle number concentration and other traffic-related air pollutants in a near-highway neighborhood over the course of a year. Atmos Environ 61: 253–264. doi: 10.1016/j.atmosenv.2012.06.088
|
| [7] |
Perkins JL, Padro-Martinez LT, Durant JL (2013) Particle number emission factors for an urban highway tunnel. Atmos Environ 74: 326–337. doi: 10.1016/j.atmosenv.2013.03.046
|
| [8] |
Ostro B, Goldberg D, Reynolds P, et al. (2015) Associations of mortality with long-term exposures to fine and ultrafine particles, species and sources: results from the California Teachers Study cohort. Environ Health Perspect 123: 549–556. doi: 10.1289/ehp.1408565
|
| [9] |
Smith SJ, Bond TC (2014) Two hundred fifty years of aerosols and climate: the end of the age of aerosols. Atmos Chem Phys 14: 537–549. doi: 10.5194/acp-14-537-2014
|
| [10] |
Bond TC (2001) Spectral dependence of visible light absorption by carbonaceous particles emitted from coal combustion. Geophys Res Lett 28: 4075–4078. doi: 10.1029/2001GL013652
|
| [11] |
Lack DA, Cappa CD (2010) Impact of brown and clear carbon on light absorption enhancement, single scatter albedo and adsorption wavelength dependance of black carbon. Atmos Chem Phys 10: 4207–4220. doi: 10.5194/acp-10-4207-2010
|
| [12] |
Smith SJ, Mizrahi A (2013) Near-term climate mitigation by short-lived forcers. Proc Natl Acad Sci 110: 14202–14206. doi: 10.1073/pnas.1308470110
|
| [13] |
Dallmann TR, Harley RA, Kirchstetter TW (2011) Effects of diesel particle filter retrofits and accelerated fleet turnover on drayage truck emissions at the Port of Oakland. Environ Sci Technol 45: 10773–10779. doi: 10.1021/es202609q
|
| [14] |
Dallmann TR, Kirchstetter TW, DeMartini SJ, et al. (2013) Quantifying on-road emissions from gasoline-powered motor vehicles: accounting for the presence of medium- and heavy-duty diesel trucks. Environ Sci Technol 47: 13873–13881. doi: 10.1021/es402875u
|
| [15] |
May AA, Nguyen NT, Presto AA, et al. (2014) Gas- and particle-phase primary emissions from in-use, on-road gasoline and diesel vehicles. Atmos Environ 88: 247–260. doi: 10.1016/j.atmosenv.2014.01.046
|
| [16] |
Robert MA, Bergen SV, Kleeman MJ, et al. (2007) Size and composition distributions of particulate matter emissions: Part 1 - light-duty gasoline vehicles. J Air Waste Manage 57: 1414– 1428. doi: 10.3155/1047-3289.57.12.1414
|
| [17] |
Robert MA, Kleeman MJ, Jakober CA (2007) Size and composition distributions of particulate matter emissions: Part 2 - heavy-duty diesel vehicles. J Air Waste Manage 57: 1429–1438. doi: 10.3155/1047-3289.57.12.1429
|
| [18] | Schauer JJ, Kleeman MJ, Cass GR, et al. (1999) Measurement of emissions from air pollution sources. 2. C2 through C30 organic compounds from medium duty diesel trucks. Environ Sci Technol 33: 1578–1587. |
| [19] | Schauer JJ, Kleeman MJ, Cass GR, et al. (2002). Measurement of emissions from air pollution sources. 5. C1 - C32 organic compounds from gasoline-powered motor vehicles. Environ Sci Technol 36: 1169–1180. |
| [20] |
Adams PJ, Seinfeld JH (2002) Predicting global aerosol size distributions in general circulation models. J Geophys Res 107: 4370 – 4392. doi: 10.1029/2001JD001010
|
| [21] |
Pierce JR, Theodoristi G, Adams PJ, et al. (2009) Parameterization of the effect of sub-grid scale aerosol dynamics on aerosol number emission rates. J Aerosol Sci 40: 285–293. doi: 10.1016/j.jaerosci.2008.11.005
|
| [22] |
Sahlodin AM, Sotudeh-Gharebagh R, Zhu Y (2007) Modeling of dispersion near roadways based on the vehicle-induced turbulence concept. Atmos Environ 41: 92–102. doi: 10.1016/j.atmosenv.2006.08.004
|
| [23] |
Wang YJ, Zhang KM (2009) Modeling near-road air quality using a computational fluid dynamics model, CFD-VIT-RIT. Environ Sci Technol 43: 7778–7783. doi: 10.1021/es9014844
|
| [24] | Zhang KM, Wexler AS (2004) Evolution of particle number distribution near roadways. part 1: analysis of aerosol dynamics and its implications for engine emission measurements. Atmos Environ 38: 6643–6653. |
| [25] | Zhang KM, Wexler AS, Zhu YF, et al. (2004) Evolution of particle number distribution near roadways. part ii: the 'road-to-ambient' process. Atmos Environ 38: 6655–6665. |
| [26] |
Zhu YF, Hinds WC, Kim S, et al. (2002) Study of ultrafine particles near a major highway with heavy-duty diesel traffic. Atmos Environ 36: 4323–4335. doi: 10.1016/S1352-2310(02)00354-0
|
| [27] |
Zhu YF, Hinds WC, Kim S, et al. (2002) Concentration and size distribution of ultrafine particles near a major highway. J Air Waste Manage Assoc 52: 1032–1042. doi: 10.1080/10473289.2002.10470842
|
| [28] |
Zhu YF, Kuhn T, Mayo P, et al. (2006) Comparison of daytime and nighttime concentration profiles and size distributions of ultrafine particles near a major highway. Environ Sci Technol 40: 2531–2536. doi: 10.1021/es0516514
|
| [29] |
Zhu YF, Hinds WC, Shen S, et al. (2004) Seasonal trends of concentration and size distribution of ultra fine particles near major highways in los angeles. Aerosol Sci Tech 38: 5–13. doi: 10.1080/02786820390229156
|
| [30] |
Huang L, Gong SL, Gordon M, et al. (2014) Aerosol-CFD modelling of ultrafine and black carbon particle emission, dilution, and growth near roadways. Atmos Chem Phys Discuss 14: 12235–12278. doi: 10.5194/acpd-14-12235-2014
|
| [31] |
Biswas S, Ntziachristos L, Moore KF, et al. (2007) Particle volatility in the vicinity of a freeway with heavy-duty diesel traffic. Atmos Environ 41: 3479–3493. doi: 10.1016/j.atmosenv.2006.11.059
|
| [32] | Salmond JA, Williams DE, Laing G, et al. (2012) The influence of vegetation on the horizontal and vertical distribution of pollutants in a street canyon. Sci Total Environ 443: 287–298. |
| [33] |
Neft I, Scungio M, Culver N, et al. (2016) Simulations of aerosol filtration by vegetation: Validation of existing models with available lab data and application to near-roadway scenario. Aerosol Sci Tech 50: 937–946. doi: 10.1080/02786826.2016.1206653
|
| [34] |
Salmond JA, Williams DE, Laing G, et al. (2013) The influence of vegetation on the horizontal and vertical distribution of pollutants in a street canyon. Sci Total Environ 443: 287–298. doi: 10.1016/j.scitotenv.2012.10.101
|
| [35] |
Scungio M, Arpino F, Stabile L, et al. (2013) Numerical simulation of ultrafine particle dispersion in urban street canyons with the spalart-allmaras turbulence model. Aerosol Air Qual Res 13: 1423–1437. doi: 10.4209/aaqr.2012.11.0306
|
| [36] |
Steffens JT, Wang YJ, Zhang KM (2012) Exploration of effects of a vegetation barrier on particle size distributions in near-road environment. Atmos Environ 50: 120–128. doi: 10.1016/j.atmosenv.2011.12.051
|
| [37] |
Steffens JT, Heist DK, Perry SG, et al. (2013) Modeling the effects of a solid barriers on pollutant dispersion under various atmospheric stability conditions. Atmos Environ 69: 76–85. doi: 10.1016/j.atmosenv.2012.11.051
|
| [38] |
Chang CH, Meroney RN (2003) Concentration and flow distributions in urban street canyons: wind tunnel and computational data. J Wind Eng Ind Aerod 91: 1141–1154. doi: 10.1016/S0167-6105(03)00056-4
|
| [39] |
Solazzo E, Cai X, Vardoulakis S (2008) Modelling wind flow and vehicle-induced turbulence in urban streets. Atmos Environ 42: 4918–4931. doi: 10.1016/j.atmosenv.2008.02.032
|
| [40] |
Hagler GSW, Tang W, Freeman MJ, et al. (2011) Model evaluation of roadside barrier impact on near-road air pollution. Atmos Environ 45: 2522–2530. doi: 10.1016/j.atmosenv.2011.02.030
|
| [41] |
Hanna SR, Tehranian S, Carissimo B, et al. (2002) Comparisons of model simulations with observations of mean flow and turbulence within simple obstacle arrays . Atmos Environ 36: 5067–5079. doi: 10.1016/S1352-2310(02)00566-6
|
| [42] |
Gowardhan AA, Pardyjak ER, Senocak I, et al. (2011) A CFD-based wind solver for an urban fast response transport and dispersion model. Environ Fluid Mech 11: 439–464. doi: 10.1007/s10652-011-9211-6
|
| [43] | Kota SH, Ying Q, Zhang Y (2013) Simulating near-road reactive dispersion of gaseous air pollutants using a three-dimensional Eulerian model. Sci Total Environ 454–455: 348–357. |
| [44] |
Wang YJ, DenBleyker A, McDonald-Buller E, et al. (2011) Modeling the chemical evolution of nitrogen oxides near roadways. Atmos Environ 45: 43–52. doi: 10.1016/j.atmosenv.2010.09.050
|
| [45] |
Wang YJ, Nguyen MT, Steffens JT, et al. (2013) Modeling multi-scale aerosol dynamics and micro-environment air quality near a large highway intersection using the ctag model. Sci Total Environ 443: 375–386. doi: 10.1016/j.scitotenv.2012.10.102
|
| [46] |
Wang YJ, Zhang KM (2012) Coupled turbulence and aerosol dynamics modeling of vehicle exhaust plumes using the CTAG model. Atmos Environ 59: 284–293. doi: 10.1016/j.atmosenv.2012.04.062
|
| [47] | Richards K, Senecal P, Pomraning E (2013) A Three-Dimensional Computational Fluid Dynamics Program for Transient or Steady State Flow with Complex Geometries. Convergent Science, Inc. Convergent Science, Inc., Middleton,WI (version 2.1.0 ed.). |
| [48] |
Issa R (1986) Solution of the implicitly discretised fluid flow equations by operator-splitting. J Comput Phys 62: 40–65. doi: 10.1016/0021-9991(86)90099-9
|
| [49] |
Tzivion S, Feingold G, Levin Z (1987) An efficient numerical solution to the stochastic collection equation. J Atmos Sci 44: 3139–3149. doi: 10.1175/1520-0469(1987)044<3139:AENSTT>2.0.CO;2
|
| [50] |
Tzivion S, Feingold G, Levin Z (1989) The evolution of raindrop spectra, part ii, collisional collection/breakup and evaporation in a rainshaft. J Atmos Sci 46: 3312–3328. doi: 10.1175/1520-0469(1989)046<3312:TEORSP>2.0.CO;2
|
| [51] |
Lee YH, Adams PJ (2012) A fast and efficient version of the two-moment aerosol sectional (TOMAS) global aerosol microphysics model. Aerosol Sci Tech 46: 678–689. doi: 10.1080/02786826.2011.643259
|
| [52] |
Shrivastava MK, Stanier CO, Robinson AL (2006) Modeling semi volatile organic aerosol mass emissions from combustion systems. Environ Sci Technol 40: 2671–2677. doi: 10.1021/es0522231
|
| [53] |
Singh S, Adams PJ, Misquitta A, et al. (2014) Computational analysis of particle nucleation in dilution tunnels: Effects of flow configuration and tunnel geometry. Aerosol Sci Tech 48: 638–648. doi: 10.1080/02786826.2014.910291
|
| [54] | Rader DJ, McMurry PH, Smith S (1987) Evaporation rates of monodispersed organic aerosols in the 0.02- to 0.2-μm-diameter range. Aerosol Sci Tech 6: 247–260. |
| [55] |
May AA, Presto AA, Hennigan CJ, et al. (2013) Gas-particle partitioning of primary organic aerosol emissions: (1) gasoline vehicle exhaust. Atmos Environ 77: 128–139. doi: 10.1016/j.atmosenv.2013.04.060
|
| [56] |
May AA, Presto AA, Hennigan CJ, et al. (2013) Gas-particle partitioning of primary organic aerosol emissions: (2) diesel vehicles. Environ Sci Technol 47: 8288–8296. doi: 10.1021/es400782j
|
| [57] |
Kalthoff N, Baumer D, Corsmeier U, et al. (2005) Vehicle-induced turbulence near a motorway. Atmos Environ 39: 5737–5749. doi: 10.1016/j.atmosenv.2004.06.048
|
| [58] |
Norris SE, Richards PJ (2011) Appropriate boundary conditions for computational wind engineering models revisited. J Wind Eng Ind Aerod 99: 257–266. doi: 10.1016/j.jweia.2010.12.008
|
| [59] | Zhang KM,Wexler AS, Niemeier DA, et al. (2005) Evolution of particle number distribution near roadways. part iii: traffic analysis and on-road size resolved particulate emission factors. Atmos Environ 39: 4155–4166. |
| [60] |
Lipsky EM, Robinson AL (2005) Design and evaluation of a portable dilution sampling system for measuring fine particle emissions from combustion systems. Aerosol Sci Tech 39: 542–553. doi: 10.1080/027868291004850
|
| [61] |
Jung JG, Pandis SN, Adams PJ (2008) Evaluation of nucleation theories in sulfur-rich environment. Aerosol Sci Technol 42: 495–504. doi: 10.1080/02786820802187085
|
| [62] |
Zhao Y, Nguyen NT, Presto AA, et al. (2015) Intermediate volatility organic compound emissions from on-road diesel vehicles: chemical composition, emission factors, and estimated secondary organic aeros production. Environ Sci Technol 49: 11516–11526. doi: 10.1021/acs.est.5b02841
|
| [63] |
Zhao Y, Nguyen NT, Presto AA, et al. (2016) Intermediate volatility organic compound emissions from on-road gasoline vehicles and small o_-road gasoline engines. Environ Sci Technol 50: 4554–4563. doi: 10.1021/acs.est.5b06247
|
| [64] |
Gkatzelis GI, Papanastasiou DK, Florou K, et al. (2016) Measurement of nonvolatile particle number size distribution. Atmos Meas Tech 9: 103–114. doi: 10.5194/amt-9-103-2016
|
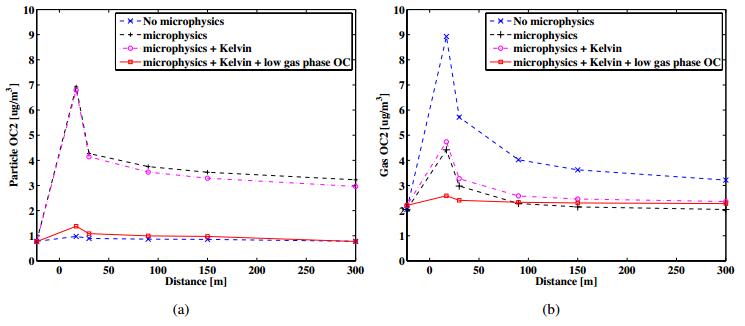
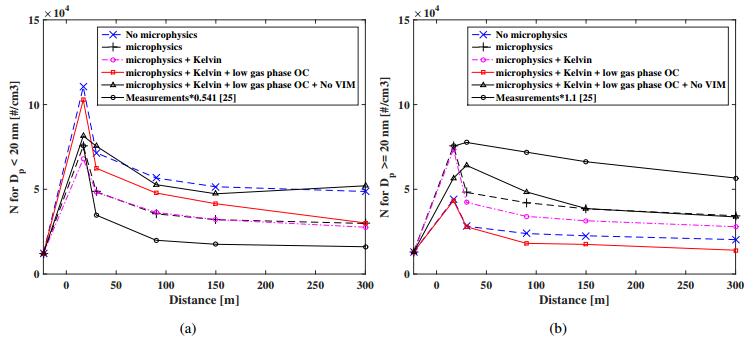
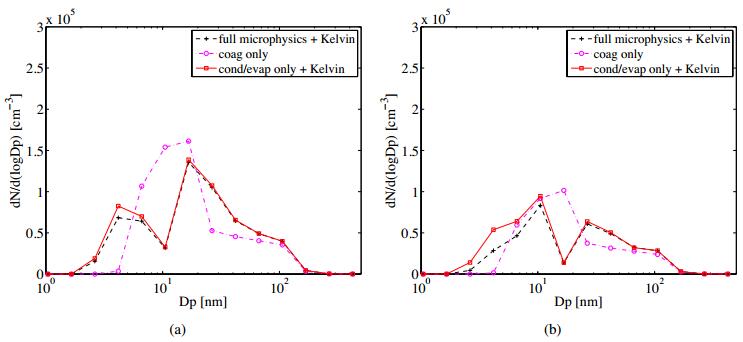
Figures(11) / Tables(2)

Satbir Singh, Peter J. Adams, Albert A. Presto. Simulations of vehicle-induced mixing and near-road aerosol microphysics using computational fluid dynamics[J]. AIMS Environmental Science, 2018, 5(5): 315-339. doi: 10.3934/environsci.2018.5.315







 DownLoad:
DownLoad: