Citation: Megan A. Ahern, Claudine P. Black, Gregory J. Seedorf, Christopher D. Baker, Douglas P. Shepherd. Hyperoxia impairs pro-angiogenic RNA production in preterm endothelial colony-forming cells[J]. AIMS Biophysics, 2017, 4(2): 284-297. doi: 10.3934/biophy.2017.2.284

| [1] |
Munsky B, Fox Z, Neuert G (2015) Integrating single-molecule experiments and discrete stochastic models to understand heterogeneous gene transcription dynamics. Methods 85: 12–21. doi: 10.1016/j.ymeth.2015.06.009

|
| [2] |
Munsky B, Neuert G, Oudenaarden AV (2012) Using gene expression noise to understand gene regulation. Science 336: 183–187. doi: 10.1126/science.1216379
|
| [3] |
Neuert G, Munsky B, Tan RZ, et al. (2013) Systematic identification of signal-activated stochastic gene regulation. Science 339: 584–587. doi: 10.1126/science.1231456
|
| [4] |
Shepherd DP, Li N, Micheva-Viteva SN, et al. (2013) Counting small RNA in pathogenic bacteria. Anal Chem 85: 4938–4943. doi: 10.1021/ac303792p
|
| [5] |
Hill JM, Zalos G, Halcox JP, et al. (2003) Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N Engl J Med 348: 593–600. doi: 10.1056/NEJMoa022287
|
| [6] |
Ingram DA, Mead LE, Tanaka H, et al. (2004) Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord. Blood 104: 2752–2760. doi: 10.1182/blood-2004-04-1396
|
| [7] |
Prater DN, Case J, Ingram DA, et al. (2007) Working hypothesis to redefine endothelial progenitor cells. Leukemia 21: 1141–1149. doi: 10.1038/sj.leu.2404676
|
| [8] |
Critser PJ, Yoder MC (2010) Endothelial colony forming cell role in neoangiogenesis and tissue repair. Curr Opin Organ Transplant 15: 68–72. doi: 10.1097/MOT.0b013e32833454b5
|
| [9] |
Fujinaga H, Baker CD, Ryan SL, et al. (2009) Hyperoxia disrupts vascular endothelial growth factor-nitric oxide signaling and decreases growth of endothelial colony-forming cells from preterm infants. Am J Physiol-Lung C 297: L1160–L1169. doi: 10.1152/ajplung.00234.2009
|
| [10] |
Baker CD, Balasubramaniam V, Mourani PM, et al. (2012) Cord blood angiogenic progenitor cells are decreased in bronchopulmonary dysplasia. Eur Respir J 40: 1516–1522. doi: 10.1183/09031936.00017312
|
| [11] | Gumina DL, Black CP, Balasubramaniam V, et al. (2016) Umbilical cord blood circulating progenitor cells and endothelial colony-forming cells are decreased in preeclampsia. Reprod Sci: 1933719116678692. |
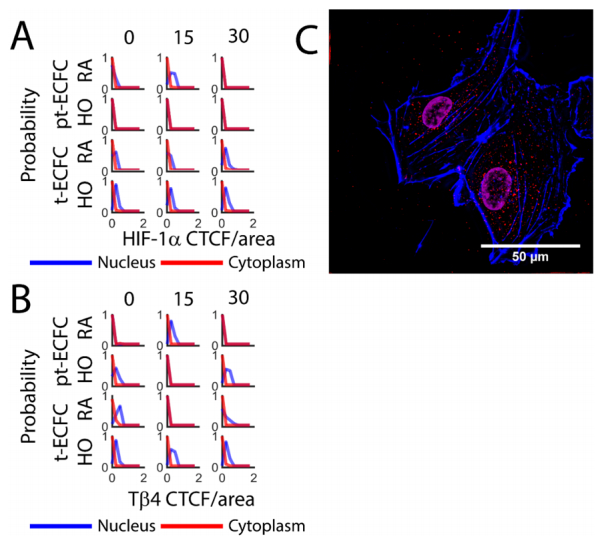
| [12] | Jo JO, Kim SR, Bae MK, et al. (2010) Thymosin β4 induces the expression of vascular endothelial growth factor (VEGF) in a hypoxia-inducible factor (HIF)-1α-dependent manner. BBA-Mol Cell Res 1803: 1244–1251. |
| [13] |
Kim NS, Kang YJ, Jo JO, et al. (2011) Elevated expression of thymosin β4, vascular endothelial growth factor (VEGF), and hypoxia inducible factor (HIF)-1α in early-stage cervical cancers. Pathol Oncol Res 17: 493–502. doi: 10.1007/s12253-010-9327-x
|
| [14] |
Moon EY, Im YS, Ryu YK, et al. (2010) Actin-sequestering protein, thymosin beta-4, is a novel hypoxia responsive regulator. Clin Exp Metastasis 27: 601–609. doi: 10.1007/s10585-010-9350-z
|
| [15] |
Oh JM, Moon EY (2010) Actin-sequestering protein, thymosin beta-4, induces paclitaxel resistance through ROS/HIF-1α stabilization in HeLa human cervical tumor cells. Life Sci 87: 286–293. doi: 10.1016/j.lfs.2010.07.002
|
| [16] |
Milosevic J, Adler I, Manaenko A, et al. (2009) Non-hypoxic stabilization of hypoxia-inducible factor alpha (HIF-α): relevance in neural progenitor/stem cells. Neurotox Res 15: 367–380. doi: 10.1007/s12640-009-9043-z
|
| [17] |
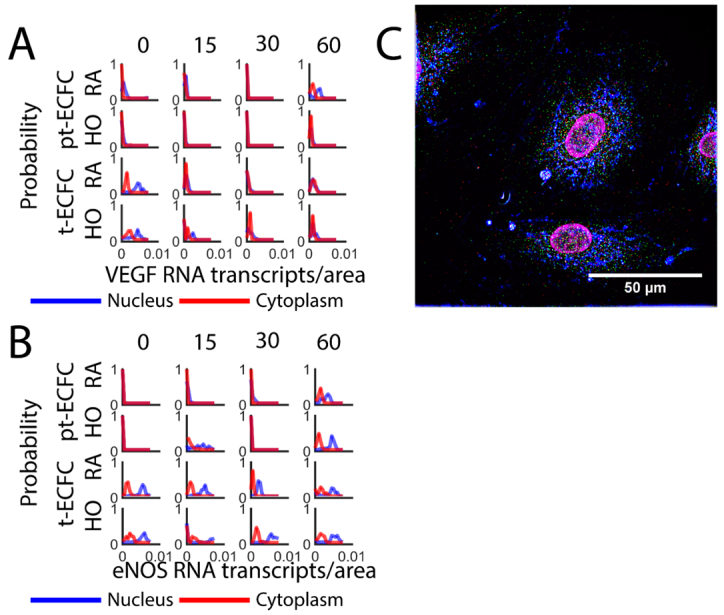
Abman SH (2010) Impaired vascular endothelial growth factor signaling in the pathogenesis of neonatal pulmonary vascular disease. Adv Exp Med Biol 661: 323–335. doi: 10.1007/978-1-60761-500-2_21
|
| [18] |
Ferrara N (2004) Vascular endothelial growth factor: basic science and clinical progress. Endocr Rev 25: 581–611. doi: 10.1210/er.2003-0027
|
| [19] |
Drummond GR, Cai H, Davis ME, et al. (2000) Transcriptional and posttranscriptional regulation of endothelial nitric oxide synthase expression by hydrogen peroxide. Circ Res 86: 347–354. doi: 10.1161/01.RES.86.3.347
|
| [20] |
Dudzinski DM, Michel T (2007) Life history of eNOS: partners and pathways. Cardiovasc Res 75: 247–260. doi: 10.1016/j.cardiores.2007.03.023
|
| [21] |
Femino AM, Fay FS, Fogarty K, et al. (1998) Visualization of single RNA transcripts in situ. Science 280: 585–590. doi: 10.1126/science.280.5363.585
|
| [22] |
Raj A, Bogaard P van den, Rifkin SA, et al. (2008) Imaging individual mRNA molecules using multiple singly labeled probes. Nat Methods 5: 877–879. doi: 10.1038/nmeth.1253
|
| [23] |
Baker CD, Ryan SL, Ingram DA, et al. (2009) Endothelial colony-forming cells from preterm infants are increased and more susceptible to hyperoxia. Am J Respir Crit Care Med 180: 454–461. doi: 10.1164/rccm.200901-0115OC
|
| [24] |
Edelstein AD, Tsuchida MA, Amodaj N, et al. (2014) Advanced methods of microscope control using μManager software. J Biol Methods 1: e10. doi: 10.14440/jbm.2014.36
|
| [25] |
Bruce MA, Butte MJ (2013) Real-time GPU-based 3D deconvolution. Opt Express 21: 4766–4773. doi: 10.1364/OE.21.004766
|
| [26] |
Mueller F, Senecal A, Tantale K, et al. (2013) FISH-Quant: automatic counting of transcripts in 3d fish images. Nat Methods 10: 277–278. doi: 10.1038/nmeth.2406
|
| [27] |
Perillo EP, De Haro L, Phipps L, et al. (2014) Enhanced 3D localization of individual RNA transcripts via astigmatic imaging. Proc SPIE 8950: 895003. doi: 10.1117/12.2038197
|
| [28] |
Székely GJ, Rizzo ML (2013) The distance correlation-test of independence in high dimension. J Multivar Anal 117: 193–213. doi: 10.1016/j.jmva.2013.02.012
|
| [29] |
Prasain N, Lee MR, Vemula S, et al. (2014) Differentiation of human pluripotent stem cells to cells similar to cord-blood endothelial colony-forming cells. Nat Biotechnol 32: 1151–1157. doi: 10.1038/nbt.3048
|
Figures(4)

Megan A. Ahern, Claudine P. Black, Gregory J. Seedorf, Christopher D. Baker, Douglas P. Shepherd. Hyperoxia impairs pro-angiogenic RNA production in preterm endothelial colony-forming cells[J]. AIMS Biophysics, 2017, 4(2): 284-297. doi: 10.3934/biophy.2017.2.284







 DownLoad:
DownLoad: