Citation: Robert Cote, Laura Lynn Eggink, J. Kenneth Hoober. CLEC receptors, endocytosis and calcium signaling[J]. AIMS Allergy and Immunology, 2017, 1(4): 207-231. doi: 10.3934/Allergy.2017.4.207

| [1] |
Das S, Dawson NL, Orengo RA (2015) Diversity in protein domain superfamilies. Curr Opin Genet Dev 35: 40–49. doi: 10.1016/j.gde.2015.09.005

|
| [2] |
Giuroiu I, Weber J (2017) Novel checkpoints and cosignaling molecules in cancer immunotherapy. Cancer J 23: 23–31. doi: 10.1097/PPO.0000000000000241
|
| [3] |
Wei SC, Levine JH, Cogdill AP, et al. (2017) Distinct cellular mechanisms underlie anti-CTLA-4 and anti-PD-1 checkpoint blockade. Cell 170: 1120–1133. doi: 10.1016/j.cell.2017.07.024
|
| [4] | Murmann AE, McMahon KM, Haluck-Kangas A, et al. (2017) Induction of DISE in ovarian cancer cells in vivo. Oncotarget 8: 84643–84658. |
| [5] |
Buchbinder E, Desai A (2016) CTLA-4 and PD-1 pathways: similarities, differences, and implications of their inhibition. Am J Clin Oncol 39: 98–106. doi: 10.1097/COC.0000000000000239
|
| [6] | Rotte A, Jin JY, Lemaire V (2017) Mechanistic overview of immune checkpoints to support the rational design of their combinations in cancer immunotherapy. Ann Oncol. |
| [7] |
Zelensky AN, Gready JE (2005) The C-type lectin-like domain superfamily. FEBS J 272: 6179–6217. doi: 10.1111/j.1742-4658.2005.05031.x
|
| [8] |
Geijtenbeek TB, Gringhuis SI (2009) Signaling through C-type lectin receptors: shaping immune responses. Nat Rev Immunol 9: 465–479. doi: 10.1038/nri2569
|
| [9] |
García-Vallejo JJ, Van KY (2009) Endogenous ligands for C-type lectin receptors: the true regulators of immune homeostasis. Immunol Rev 230: 22–37. doi: 10.1111/j.1600-065X.2009.00786.x
|
| [10] |
Van KY, Ilarregui JM, van Vliet SJ (2015) Novel insights into the immunomodulatory role of the dendritic cell and macrophage-expressed C-type lectin MGL. Immunobiology 220: 185–192. doi: 10.1016/j.imbio.2014.10.002
|
| [11] |
Sancho D, Reis SC (2013) Sensing of cell death by myeloid C-type lectin receptors. Curr Opin Immunol 25: 46–52. doi: 10.1016/j.coi.2012.12.007
|
| [12] |
Chang SY, Kweon MN (2010) Langerin-expressing dendritic cells in gut-associated lymphoid tissues. Immunol Rev 234: 233–246. doi: 10.1111/j.0105-2896.2009.00878.x
|
| [13] |
Ingeborg SO, Unger WWJ, Yvette VK (2011) C-type lectin receptors for tumor eradication: future directions. Cancers 3: 3169–3188. doi: 10.3390/cancers3033169
|
| [14] |
Zhang F, Ren S, Zuo Y (2014) DC-SIGN, DC-SIGNR and LSECtin: C-type lectins for infection. Int Rev Immunol 33: 54–66. doi: 10.3109/08830185.2013.834897
|
| [15] | Yan H, Kamiya T, Suabjakyong P, et al. (2015) Targeting C-type lectin receptors for cancer immunity. Front Immunol 6: 408–416. |
| [16] |
Dambuza IM, Brown GD (2015) C-type lectins in immunity: recent developments. Curr Opin Immunol 32: 21–27. doi: 10.1016/j.coi.2014.12.002
|
| [17] | Ding D, Yao Y, Zhang S, et al. (2017) C-type lectins facilitate tumor metastasis. Oncol Lett 13: 13–21. |
| [18] |
Andersen CBF, Moestrup SK (2014) How calcium makes endocytic receptors attractive. Trends Biochem Sci 39: 82–90. doi: 10.1016/j.tibs.2013.12.003
|
| [19] | Eggensperger A, Tampé R (2015) The transporter associated with antigen processing: a key player in adaptive immunity. Biol Chem 396: 1059–1072. |
| [20] |
Rodriguez A, Regnault A, Kleijmeer M, et al. (1999) Selective transport of internalized antigens to the cytosol for MHC class I presentation in dendritic cells. Nat Cell Biol 1: 362–368. doi: 10.1038/14058
|
| [21] |
Solheim JC (1999) Class I MHC molecules: assembly and antigen presentation. Immunol Rev 172: 11–19. doi: 10.1111/j.1600-065X.1999.tb01352.x
|
| [22] |
Gil-Torregrosa BC, Lennon-Duménil AM, Kessler B, et al. (2004) Control of cross-presentation during dendritic cell maturation. Eur J Immunol 34: 398–407. doi: 10.1002/eji.200324508
|
| [23] |
Heath WR, Carbone FR (2001) Cross-presentation in viral immunity and self-tolerance. Nat Rev Immunol 1: 126–135. doi: 10.1038/35100512
|
| [24] |
Joffre OP, Segura E, Savina A, et al. (2012) Cross-presentation by dendritic cells. Nat Rev Immunol 12: 557–569. doi: 10.1038/nri3254
|
| [25] | McDonnell AM, Robinson BWS, Currie AJ (2010) Tumor antigen cross-presentation and the dendritic cell: where it all begins? Clin Dev Immunol 2010: 539519–539527. |
| [26] | Bousso P, Robey E (2003) Dynamics of CD8+ T cell priming by dendritic cells in intact lymph nodes. Nat Immunol 4: 579–585. |
| [27] |
Randolph GJ, Jakubzick C, Qu C (2008) Antigen presentation by monocytes and monocyte-derived cells. Curr Opin Immunol 20: 52–60. doi: 10.1016/j.coi.2007.10.010
|
| [28] |
Leiri?o P, Fresno CD, Ardavín C (2012) Monocytes as effector cells: activated Ly-6C(high) mouse monocytes migrate to the lymph nodes through the lymph and cross-present antigens to CD8+ T cells. Eur J Immunol 42: 2042–2051. doi: 10.1002/eji.201142166
|
| [29] |
Raghavan M, Wijeyesakere SJ, Peters LR, et al. (2013) Calreticulin in the immune system: ins and outs. Trends Immunol 34: 13–21. doi: 10.1016/j.it.2012.08.002
|
| [30] |
Lv D, Shen Y, Peng Y, et al. (2015) Neuronal MHC class I expression is regulated by activity driven calcium signaling. PLoS One 10: e0135223–e0135238. doi: 10.1371/journal.pone.0135223
|
| [31] | Skov S (1999) Intracellular signal transduction mediated by ligation of MHC class I molecules. Tissue Antigens 51: 215–223. |
| [32] |
Blum JS, Wearsch PA, Cresswell P (2013) Pathways of antigen processing. Annu Rev Immunol 31: 443–473. doi: 10.1146/annurev-immunol-032712-095910
|
| [33] |
Roche PA, Furuta K (2015) The ins and outs of MHC class II-mediated antigen processing and presentation. Nat Rev Immunol 15: 203–216. doi: 10.1038/nri3818
|
| [34] |
Mueller SN (2017) Spreading the load: antigen transfer between migratory and lymph node-resident dendritic cells promotes T-cell priming. Eur J Immunol 47: 1798–1801. doi: 10.1002/eji.201747248
|
| [35] |
Parton RG, Joggerst B, Simons K (1994) Regulated internalization of caveolae. J Cell Biol 127: 1199–1215. doi: 10.1083/jcb.127.5.1199
|
| [36] |
Levine TP, Chain BM (1992) Endocytosis by antigen presenting cells: dendritic cells are as endocytically active as other antigen presenting cells. Proc Natl Acad Sci USA 89: 8342–8346. doi: 10.1073/pnas.89.17.8342
|
| [37] |
Hohn C, Lee SR, Pinchuk LM, et al. (2009) Zebrafish kidney phagocytes utilize macropinocytosis and Ca+-dependent endocytic mechanisms. PLoS One 4: e4314–e4323. doi: 10.1371/journal.pone.0004314
|
| [38] |
Calmette J, Bertrand M, Vétillard M, et al. (2016) Glucocorticoid-induced leucine zipper protein controls macropinocytosis in dendritic cells. J Immunol 197: 4247–4256. doi: 10.4049/jimmunol.1600561
|
| [39] |
Ayroldi E, Riccardi C (2009) Glucocorticoid-induced leucine zipper (GILZ): a new important mediator of glucocorticoid action. FASEB J 23: 3649–3658. doi: 10.1096/fj.09-134684
|
| [40] | Ronchetti S, Migliorati G, Riccardi C (2015) GILZ as a mediator of the anti-inflammatory effects of glucocorticoids. Front Endocrinol 6: 170–175. |
| [41] |
Canton J, Schlam D, Breuer C, et al. (2016) Calcium-sensing receptors signal constitutive macropinocytosis and facilitate the uptake of NOD2 ligands in macrophages. Nat Commun 7: 11284–11295. doi: 10.1038/ncomms11284
|
| [42] | Redka DS, Gütschow M, Grinstein S, et al. (2017) Differential ability of pro-inflammatory and anti-inflammatory macrophages to perform macropinocytosis. Mol Biol Cell pii: mbc.E17-06-0419. |
| [43] | Carafoli E, Krebs J (2016) Why calcium? How calcium became the best communicator. J Biol Chem 291: 20849–20857. |
| [44] |
Caroppo R, Gerbino A, Fistetto G, et al. (2004) Extracellular calcium acts as a "third messenger" to regulate enzyme and alkaline secretion. J Cell Biol 166: 111–119. doi: 10.1083/jcb.200310145
|
| [45] | Drickamer K (1988) Two distinct classes of carbohydrate-recognition domains in animal lectins. J Biol Chem 263: 9557–9560. |
| [46] |
Drickamer K (1996) Ca(2+)-dependent sugar recognition by animal lectins. Biochem Soc T 24: 146–150. doi: 10.1042/bst0240146
|
| [47] |
Weis WI, Taylor ME, Drickamer K (1998) The C-type lectin superfamily in the immune system. Immunol Rev 163: 19–34. doi: 10.1111/j.1600-065X.1998.tb01185.x
|
| [48] |
Drickamer K, Taylor ME (2015) Recent insights into structures and functions of C-type lectins in the immune system. Curr Opin Struc Biol 34: 26–34. doi: 10.1016/j.sbi.2015.06.003
|
| [49] |
van Vliet SJ, Saeland E, Van KY (2008) Sweet preferences of MGL: carbohydrate specificity and function. Trends Immunol 29: 83–90. doi: 10.1016/j.it.2007.10.010
|
| [50] |
Van DD, Stolk DA, Van RVD, et al. (2017) Targeting C-type lectin receptors: a high-carbohydrate diet for dendritic cells to improve cancer vaccines. J Leukocyte Biol 102: 1017–1034. doi: 10.1189/jlb.5MR0217-059RR
|
| [51] |
Napoletano C, Zizzari IG, Rughetti A, et al. (2012) Targeting of macrophage galactose-type C-type lectin (MGL) induces DC signaling and activation. Eur J Immunol 42: 936–945. doi: 10.1002/eji.201142086
|
| [52] |
Engering A, Geijtenbeck TBH, van Vliet SJ, et al. (2002) The dendritic cell-specific adhesion receptor DC-SIGN internalizes antigen for presentation to T cells. J Immunol 168: 2118–2126. doi: 10.4049/jimmunol.168.5.2118
|
| [53] | Database of human proteins containing CTLDs. Available from: http://www.imperial.ac.uk/research/animallectins/ctld/mammals/humandata%20updated.html. |
| [54] | Cummings RD, McEver RP (2017) Chapter 34: C-Type lectins, In: Varki A, Cummings RD, Esko JD, et al., editors. Essentials of Glycobiology 3rd Ed. Cold Spring Harbor Laboratory Press, 2015–2017. |
| [55] |
Sancho D, Reis SC (2012) Signaling by myeloid C-type lectin receptors in immunity and homeostasis. Annu Rev Immunol 30: 491–529. doi: 10.1146/annurev-immunol-031210-101352
|
| [56] |
Billadeau DD, Leibson PJ (2002) ITAMs versus ITIMs: striking a balance during cell regulation. J Clin Invest 109: 161–168. doi: 10.1172/JCI0214843
|
| [57] |
Ivashkiv LB (2009) Cross-regulation of signaling by ITAM-associated receptors. Nat Immunol 10: 340–347. doi: 10.1038/ni.1706
|
| [58] |
Bezbradica JS, Rosenstein RK, DeMarco RA, et al. (2014) A role for the ITAM signaling module in specifying cytokine-receptor functions. Nat Immunol 15: 333–342. doi: 10.1038/ni.2845
|
| [59] |
Pollitt AY, Poulter NS, Gitz E, et al. (2014) Syk and Src family kinases regulate C-type lectin receptor 2 (CLEC-2)-mediated clustering of podoplanin and platelet adhesion to lymphatic endothelial cells. J Biol Chem 289: 35695–35710. doi: 10.1074/jbc.M114.584284
|
| [60] |
Unkeless JC, Jin J (1997) Inhibitory receptors, ITIM sequences and phosphatases. Curr Opin Immunol 9: 338–343. doi: 10.1016/S0952-7915(97)80079-9
|
| [61] |
van Vliet SJ, Aarnoudse CA, Vc BDB, et al. (2007) MGL-mediated internalization and antigen presentation by dendritic cells: a role for tyrosine-5. Eur J Immunol 37: 2075–2081. doi: 10.1002/eji.200636838
|
| [62] | Harris RL, Cw VDB, Bowen DJ (2012) ASGR1 and ASGR2, the genes that encode the asialoglycoprotein receptor (Ashwell Receptor), are expressed in peripheral blood monocytes and show inter-individual differences in transcript profile. Mol Biol Int 2012: 283974–283983. |
| [63] |
East L, Isacke CM (2002) The mannose receptor family. BBA-Gen Subjects 1572: 364–386. doi: 10.1016/S0304-4165(02)00319-7
|
| [64] | Uniport. Available from: http://www.uniprot.org/uniprot/P22897. |
| [65] |
Lo YL, Liou GG, Lyu JH, et al. (2016) Dengue virus infection is through a cooperative interaction between a mannose receptor and CLEC5A on macrophage as a multivalent hetero-complex. PLoS One 11: e0166474–e0166486. doi: 10.1371/journal.pone.0166474
|
| [66] | R?dgaard-Hansen S, Rafique A, Christensen PA, et al. (2014) A soluble form of the macrophage-related mannose receptor (MR/CD206) is present in human serum and elevated in critical illness. Clin Chem Lab Med 52: 453–461. |
| [67] |
Feinberg H, Park-Snyder S, Kolatkar AR, et al. (2000) Structure of a C-type carbohydrate recognition domain from the macrophage mannose receptor. J Biol Chem 275: 21539–21548. doi: 10.1074/jbc.M002366200
|
| [68] |
Ng KKS, Park-Snyder S, Weis WI (1998) Ca2+-dependent structural changes in C-type mannose-binding proteins. Biochemistry 37: 17965–17976. doi: 10.1021/bi981972a
|
| [69] | Iobst ST, Wormald MR, Weis WI, et al. (1994) Binding of sugar ligands to Ca(2+)-dependent animal lectins. I. Analysis of mannose binding by site-directed mutagenesis and NMR. J Biol Chem 269: 15505–15511. |
| [70] |
Weis WI, Drickamer K, Hendrickson WA (1992) Structure of a C-type mannose-binding protein complexed with an oligosaccharide. Nature 360: 127–134. doi: 10.1038/360127a0
|
| [71] |
Drickamer K (1992) Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 360: 183–186. doi: 10.1038/360183a0
|
| [72] |
Apostolopoulos V, Pietersz GA, Loveland, BE, et al. (1995) Oxidative/reductive conjugation of mannan to antigen selects for T1 or T2 immune responses. Proc Natl Acad Sci USA 92: 10128–10132. doi: 10.1073/pnas.92.22.10128
|
| [73] |
Apostolopoulos V, Pietersz GA, Gordon S, et al. (2000) Aldehyde-mannan antigen complexes target the MHC class I antigen-presentation pathway. Eur J Immunol 30: 1714–1723. doi: 10.1002/1521-4141(200006)30:6<1714::AID-IMMU1714>3.0.CO;2-C
|
| [74] |
Apostolopoulos V, Pietersz GA, Tsibanis A, et al. (2014) Dendritic cell immunotherapy: clinical outcomes. Clin Transl Immunol 3: e21–e24. doi: 10.1038/cti.2014.14
|
| [75] |
Steinman RM, Turley S, Mellman I, et al. (2000) The induction of tolerance by dendritic cells that have captured apoptotic cells. J Exp Med 191: 411–416. doi: 10.1084/jem.191.3.411
|
| [76] |
Steinman RM, Hawiger D, Liu K, et al. (2003) Dendritic cell function in vivo during the steady state: a role in peripheral tolerance. Ann Ny Acad Sci 987: 15–25. doi: 10.1111/j.1749-6632.2003.tb06029.x
|
| [77] |
Redmond WL, Sherman LA (2005) Peripheral tolerance of CD8 T lymphocytes. Immunity 22: 275–284. doi: 10.1016/j.immuni.2005.01.010
|
| [78] |
Chieppa M, Bianchi G, Doni A, et al. (2003) Cross-linking of the mannose receptor on monocyte-derived dendritic cells activates an anti-inflammatory immunosuppressive program. J Immunol 171: 4552–4560. doi: 10.4049/jimmunol.171.9.4552
|
| [79] | Allavena P, Chieppa M, Blanchi G, et al. (2010) Engagement of the mannose receptor by tumoral mucins activates an immune suppressive phenotype in human tumor-associated macrophages. Clin Dev Immunol 2010: 547179–547188. |
| [80] |
Sharpe AH (2009) Mechanisms of costimulation. Immunol Rev 229: 5–11. doi: 10.1111/j.1600-065X.2009.00784.x
|
| [81] |
Chen L, Flies DB (2013) Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol 13: 227–242. doi: 10.1038/nri3405
|
| [82] |
Anderson PJ, Kokame K, Sadler JE (2006) Zinc and calcium ions cooperatively modulate ADAMTS13 activity. J Biol Chem 281: 850–857. doi: 10.1074/jbc.M504540200
|
| [83] | Sorvillo N, Pos W, Lm VDB, et al. (2017) The macrophage mannose receptor promotes uptake of ADAMTS13 by dendritic cells. Blood 119: 3828–3835. |
| [84] |
Mahnke K, Guo M, Lee S, et al. (2000) The dendritic cell receptor for endocytosis, DEC-205, can recycle and enhance antigen presentation via major histocompatibility complex class II-positive lysosomal compartments. J Cell Biol 151: 673–683. doi: 10.1083/jcb.151.3.673
|
| [85] |
Platt CD, Ma JK, Chalouni C, et al. (2010) Mature dendritic cells use endocytic receptors to capture and present antigens. Proc Nat Acad Sci USA 107: 4287–4292. doi: 10.1073/pnas.0910609107
|
| [86] |
Tel J, Benitez-Ribas D, Hoosemans S, et al. (2011) DEC-205 mediates antigen uptake and presentation by both resting and activated human plasmacytoid dendritic cells. Eur J Immunol 41: 1014–1023. doi: 10.1002/eji.201040790
|
| [87] |
Hawiger D, Inaba K, Dorsett Y, et al. (2001) Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J Exp Med 194: 769–779. doi: 10.1084/jem.194.6.769
|
| [88] |
Ma DY, Clark EA (2009) The role of CD40 and CD154/CD40L in dendritic cells. Semin Immunol 21: 265–272. doi: 10.1016/j.smim.2009.05.010
|
| [89] |
Lee GH, Askari A, Malietzis G, et al. (2014) The role of CD40 expression in dendritic cell in cancer biology: a systematic review. Curr Cancer Drug Tar 14: 610–620. doi: 10.2174/1568009614666140828103253
|
| [90] |
Sartorius R, D'Apice L, Trovato M, et al. (2015) Antigen delivery by filamentous bacteriophage fd displaying an anti-DEC-205 single-chain variable fragment confers adjuvanticity by triggering a TLR9-mediated immune response. Embo Mol Med 7: 973–988. doi: 10.15252/emmm.201404525
|
| [91] |
Melander MC, Jürgensen HJ, Madsen DH, et al. (2015) The collagen receptor uPARAP/Endo180 in tissue degradation and cancer (Review). Int J Oncol 47: 1177–1188. doi: 10.3892/ijo.2015.3120
|
| [92] |
Sturge J (2016) Endo180 at the cutting edge of bone cancer treatment and beyond. J Pathol 238: 485–488. doi: 10.1002/path.4673
|
| [93] |
Yuan C, Jürgensen HJ, Engelholm LH, et al. (2016) Crystal structures of the ligand-binding region of uPARAP: effect of calcium ion binding. Biochem J 473: 2359–2368. doi: 10.1042/BCJ20160276
|
| [94] |
East L, Rushton S, Taylor ME, et al. (2002) Characterization of sugar binding by the mannose receptor family member, Endo180. J Biol Chem 277: 50469–50475. doi: 10.1074/jbc.M208985200
|
| [95] |
Augert A, Payré C, de Launoit Y, et al. (2009) The M-type receptor PLA2R regulates senescence through the p53 pathway. Embo Rep 10: 271–277. doi: 10.1038/embor.2008.255
|
| [96] |
Jr BLH, Bonegio RG, Lambeau G, et al. (2009) M-type phospholipase A2 receptor as target antigen in idiopathic membranous nephropathy. New Engl J Med 361: 11–21. doi: 10.1056/NEJMoa0810457
|
| [97] |
Takahashi S, Watanabe K, Watanabe Y, et al. (2015) C-type lectin-like domain and fibronectin-like type II domain of phospholipase A2 receptor 1 modulate binding and migratory responses to collagen. Febs Lett 589: 829–835. doi: 10.1016/j.febslet.2015.02.016
|
| [98] |
Nolin JD, Ogden HL, Lai Y, et al. (2016) Identification of epithelial phospholipase A2 receptor 1 as a potential target in asthma. Am J Resp Cell Mol 55: 825–836. doi: 10.1165/rcmb.2015-0150OC
|
| [99] |
Fresquet M, Jowitt TA, McKenzie EA, et al. (2017) PLA2R binds to the annexin A2-S100A10 complex in human podocytes. Sci Rep 7: 6876–6886. doi: 10.1038/s41598-017-07028-8
|
| [100] |
Santamaria-Kisiel L, Rintala-Dempsey A, Shaw GS (2006) Calcium-dependent and -independent interactions of the S100 protein family. Biochem J 396: 201–214. doi: 10.1042/BJ20060195
|
| [101] |
Goder V, Spiess M (2001) Topogenesis of membrane proteins: determinants and dynamics. Febs Lett 504: 87–93. doi: 10.1016/S0014-5793(01)02712-0
|
| [102] |
Zimmerman R, Eyrisch S, Ahmad M, et al. (2011) Protein translocation across the ER membrane. BBA-Biomembranes 1808: 912–924. doi: 10.1016/j.bbamem.2010.06.015
|
| [103] |
Feinberg H, Mitchell DA, Drickamer K, et al. (2001) Structural basis for selective recognition of oligosaccharides by DC-SIGN and DC-SIGNR. Science 294: 2163–2166. doi: 10.1126/science.1066371
|
| [104] | Mitchell DA, Fadden AJ, Drickamer K (2001) A novel mechanism of carbohydrate recognition by the C-type lectins DC-SIGN and DC-SIGNR. Subunit organization and binding to multivalent ligands. J Biol Chem 276: 28939–28945. |
| [105] |
Guo Y, Feinberg H, Conroy E, et al. (2004) Structural basis for distinct ligand-binding and targeting properties of the receptors DC-SIGN and DC-SIGNR. Nat Struct Mol Biol 11: 591–598. doi: 10.1038/nsmb784
|
| [106] |
Caparrós E, Munoz P, Sierra-Filardi E, et al. (2006) DC-SIGN ligation on dendritic cells results in ERK and PI3K activation and modulates cytokine production. Blood 107: 3950–3958. doi: 10.1182/blood-2005-03-1252
|
| [107] |
Iyori M, Ohtani M, Hasebe A, et al. (2008) A role of the Ca2+ binding site of DC-SIGN in the phagocytosis of E. coli. Biochem Bioph Res Co 377: 367–372. doi: 10.1016/j.bbrc.2008.09.142
|
| [108] |
Dos Santos á, Hadjivasiliou A, Ossa F, et al. (2017) Oligomerization domains in the glycan-binding receptors DC-SIGN and DC-SIGNR: sequence variation and stability differences. Protein Sci 26: 306–316. doi: 10.1002/pro.3083
|
| [109] |
Dodagatta-Marri E, Mitchell DA, Pandit H, et al. (2017) Protein-protein interaction between surfactant protein D and DC-SIGN via C-type lectin domain can suppress HIV-1 transfer. Front Immunol 8: 834–845. doi: 10.3389/fimmu.2017.00834
|
| [110] |
Chao PZ, Hsieh MS, Cheng CW, et al. (2015) Dendritic cells respond to nasopharygeal carcinoma cells through annexin A2-recognizing DC-SIGN. Oncotarget 6: 159–170. doi: 10.18632/oncotarget.2700
|
| [111] |
Chia J, Goh G, Bard F (2016) Short O-GalNAc glycans: regulation and role in tumor development and clinical perspectives. BBA-Gen Subjects 1860: 1623–1639. doi: 10.1016/j.bbagen.2016.03.008
|
| [112] |
Zheng J, Xiao H, Wu R (2017) Specific identification of glycoproteins bearing the Tn antigen in human cells. Angew Chem 129: 7213–7217. doi: 10.1002/ange.201702191
|
| [113] |
Feinberg H, Torgersen D, Drickamer K, et al. (2000) Mechanism of pH-dependent N-acetylgalactosamine binding by a functional mimic of the hepatocyte asialoglycoprotein receptor. J Biol Chem 275: 35176–35184. doi: 10.1074/jbc.M005557200
|
| [114] | Morell AG, Gregoriadis G, Scheinberg IH, et al. (1971) The role of sialic acid in determining the survival of glycoproteins in the circulation. J Biol Chem 246: 1461–1467. |
| [115] |
Grewal PK (2010) The Ashwell-Morell receptor. Method Enzymol 479: 223–241. doi: 10.1016/S0076-6879(10)79013-3
|
| [116] |
Weigel PH, Yik JHN (2002) Glycans as endocytosis signals: the cases of the asialoprotein and hyaluronan/chrondroitin sulfate receptors. BBA-Gen Subjects 1572: 341–363. doi: 10.1016/S0304-4165(02)00318-5
|
| [117] | Dixon LJ, Barnes M, Tang H, et al. (2013) Kupffer Cells in the Liver. Compr Physiol 3: 785–797. |
| [118] |
Tsuiji M, Fujimori M, Ohashi Y, et al. (2002) Molecular cloning and characterization of a novel mouse macrophage C-type lectin, mMGL2, which has a distinct carbohydrate specificity from mMGL1. J Biol Chem 277: 28892–28901. doi: 10.1074/jbc.M203774200
|
| [119] |
Kolatkar AR, Weis WI (1996) Structural basis of galactose recognition by C-type animal lectins. J Biol Chem 271: 6679–6685. doi: 10.1074/jbc.271.12.6679
|
| [120] |
Meier M, Bider MD, Malashkevich VN, et al. (2000) Crystal structure of the carbohydrate recognition domain of the H1 subunit of the asialoglycoprotein receptor. J Mol Biol 300: 857–865. doi: 10.1006/jmbi.2000.3853
|
| [121] |
Higashi N, Fujioka K, Denda-Nagai K, et al. (2002) The macrophage C-type lectin specific for galactose/N-acetylgalactosamine is an endocytic receptor expressed on monocyte-derived immature dendritic cells. J Biol Chem 277: 20686–20693. doi: 10.1074/jbc.M202104200
|
| [122] |
Lundberg K, Rydnert F, Broos S, et al. (2016) C-type lectin receptor expression on human basophils and effects of allergen-specific immunotherapy. Scand J Immunol 84: 150–157. doi: 10.1111/sji.12457
|
| [123] |
Vukman KV, Ravidà A, Aldridge AM, et al. (2013) Mannose receptor and macrophage galactose-type lectin are involved in Bordetella pertussis mast cell interaction. J Leukocyte Biol 94: 439–448. doi: 10.1189/jlb.0313130
|
| [124] |
Savola P, Kelkka T, Rajala HL, et al. (2017) Somatic mutations in clonally expanded cytotoxic T lymphocytes in patients with newly diagnosed rheumatoid arthritis. Nat Commun 8: 15869–15882. doi: 10.1038/ncomms15869
|
| [125] | Gaur P, Myles A, Misra R, et al. (2016) Intermediate monocytes are increased in enthesitis-related arthritis, a category of juvenile idiopathic arthritis. Clin Exp Immunol 187: 234–241. |
| [126] |
Vlismas A, Bletsa R, Mavrogianni D, et al. (2016) Microarray analyses reveal marked differences in growth factor and receptor expression between 8-cell human embryos and pluripotent stem cells. Stem Cells Dev 25: 160–177. doi: 10.1089/scd.2015.0284
|
| [127] |
Klimmeck D, Hanssong J, Raffel S, et al. (2012) Proteomic cornerstones of hematopoietic stem cell differentiation: distinct signatures of multipotent progenitors and myeloid committed cells. Molec Cell Proteomics 11: 286–302. doi: 10.1074/mcp.M111.016790
|
| [128] |
Winkler C, Witte L, Moraw N, et al. (2014) Impact of endobronchial allergen provocation on macrophage phenotype in asthmatics. BMC Immunol 15: 12–22. doi: 10.1186/1471-2172-15-12
|
| [129] |
Mathews JA, Kasahara DI, Ribeiro L, et al. (2015) γδ T cells are required for M2 macrophage polarization and resolution of ozone-induced pulmonary inflammation in mice. PLoS One 10: e0131236–e0131251. doi: 10.1371/journal.pone.0131236
|
| [130] |
Shin H, Kumamoto Y, Gopinath S, et al. (2016) CD301b+ dendritic cells stimulate tissue-resident memory CD8+ T cells to protect against genital HSV-2. Nat Commun 7: 13346–13355. doi: 10.1038/ncomms13346
|
| [131] |
Linehan JL, Dileepan T, Kashem SW, et al. (2015) Generation of Th17 cells in response to intranasal infection requires TGF-β1 from dendritic cells and IL-6 from CD301b+ dendritic cells. Proc Natl Acad Sci USA 112: 12782–12787. 132. Wong KL, Tai JJ, Wong WC, et al. (2011) Gene expression profiling reveals the defining features of the classical, intermediate, and nonclassical human monocyte subsets. Blood 118: e16–e31. doi: 10.1182/blood-2010-12-326355
|
| [132] |
133. Wong KL, Yeap WH, Tai JJY, et al. (2012) The three human monocyte subsets: implications for health and disease. Immunol Res 53: 41–57. doi: 10.1007/s12026-012-8297-3
|
| [133] |
134. Michlmayr D, Andrade P, Gonzalez K, et al. (2017) CD14+CD16+ monocytes are the main target of Zika virus infection in peripheral blood mononuclear cells in a paediatric study in Nicaragua. Nat Microbiol 2: 1462–1470. doi: 10.1038/s41564-017-0035-0
|
| [134] |
135. Knudsen NH, Lee CH (2016) Identity crisis: CD301b(+) mononuclear phagocytes blur the M1-M2 macrophage line. Immunity 45: 461–463. doi: 10.1016/j.immuni.2016.09.004
|
| [135] |
136. Zhang W, Xu W, Xiong S (2011) Macrophage differentiation and polarization via phosphatidylinositol 3-kinase/Akt-ERK signaling pathway conferred by serum amyloid P component. J Immunol 187: 1764–1777. doi: 10.4049/jimmunol.1002315
|
| [136] |
137. Trowbridge IS, Thomas M (1994) CD45: an emerging role as a protein tyrosine phosphatase required for lymphocyte activation and development. Annu Rev Immunol 12: 85–116. doi: 10.1146/annurev.iy.12.040194.000505
|
| [137] | 138. van Vliet SJ, Gringhuis SI, Geijtenbeek TBH, et al. (2006) Regulation of effector T cells by antigen-presenting cells via interaction with the C-type lectin MGL with CD45. Nat Immunol 11: 1200–1208. |
| [138] |
139. Nam HJ, Poy F, Saito H, et al. (2005) Structural basis for the function and regulation of the receptor protein tyrosine phosphatase CD45. J Exp Med 201: 441–452. doi: 10.1084/jem.20041890
|
| [139] |
140. Xu Z, Weiss A (2002) Negative regulation of CD45 by differential homodimerization of the alternatively spliced isoforms. Nat Immunol 3: 764–771. doi: 10.1038/ni822
|
| [140] |
141. Kumar V, Cheng P, Condamine T, et al. (2016) CD45 phosphatase inhibits STAT3 transcription factor activity in myeloid cells and promotes tumor-associated macrophage differentiation. Immunity 44: 303–315. doi: 10.1016/j.immuni.2016.01.014
|
| [141] |
142. van Vliet SJ, van Liempt E, Geijtenbeek TB, et al. (2006) Differential regulation of C-type lectin expression on tolerogenic dendritic cell subsets. Immunobiology 211: 577–585. doi: 10.1016/j.imbio.2006.05.022
|
| [142] |
143. Marcelo F, Garcia-Martin F, Matsushita T, et al. (2014) Delineating binding modes of Gal/GalNAc and structural elements of the molecular recognition of tumor-associated mucin glycopeptides by the human macrophage galactose-type lectin. Chem Eur J 20: 16147–16155. doi: 10.1002/chem.201404566
|
| [143] |
144. Tanaka J, Gleinich AS, Zhang Q, et al. (2017) Specific and differential binding of N-acetylgalactosamine glycopolymers to the human macrophage galactose lectin and asialoglycoprotein receptor. Biomacromolecules 18: 1624–1633. doi: 10.1021/acs.biomac.7b00228
|
| [144] |
145. Khorev O, Stokmaier D, Schwardt O, et al. (2008) Trivalent, Gal/GaNAc-containing ligands designed for the asialoglycoprotein receptor. Bioorg Med Chem 16: 5216–5231. doi: 10.1016/j.bmc.2008.03.017
|
| [145] |
146. Nair JK, Willoughby JLS, Chan A, et al. (2014) Multivalent N-acetylgalactosamine-conjugated siRNA localizes in hepatocytes and elicits robust RNAi-mediated gene silencing. J Am Chem Soc 136: 16958–16961. doi: 10.1021/ja505986a
|
| [146] | 147. Lo-Man R, Bay S, Vichier-Guerre S, et al. (1999) A fully synthetic immunogen carrying a carcinoma-associated carbohydrate for active specific immunotherapy. Cancer Res 59: 1520–1524. |
| [147] |
148. Lo-Man R, Vichier-Guerre S, Bay S, et al. (2001) Anti-tumor immunity provided by a synthetic multiple antigenic glycopeptide displaying a tri-Tn glycotope. J Immunol 166: 2849–2854. doi: 10.4049/jimmunol.166.4.2849
|
| [148] |
149. Morgan AJ, Platt FM, Lloyd-Evans E, et al. (2011) Molecular mechanisms of endolysosomal Ca2+ signaling in health and disease. Biochem J 439: 349–374. doi: 10.1042/BJ20110949
|
| [149] |
150. Wragg S, Drickamer K (1999) Identification of amino acid residues that determine pH dependence of ligand binding to the asialoglyprotein receptor during endocytosis. J Biol Chem 274: 35400–35406. doi: 10.1074/jbc.274.50.35400
|
| [150] |
151. Onizuka T, Shimizu H, Moriwaki Y, et al. (2012) NMR study of ligand release from asialoglycoprotein receptor under solution conditions in early endosomes. Febs J 279: 2645–2656. doi: 10.1111/j.1742-4658.2012.08643.x
|
| [151] |
152. Gerasimenko JV, Tepikin AV, Petersen OH, et al. (1998) Calcium uptake via endocytosis with rapid release from acidifying endosomes. Curr Biol 8: 1335–1338. doi: 10.1016/S0960-9822(07)00565-9
|
| [152] |
153. Plattner H, Verkhratsky A (2016) Inseparable tandem: evolution chooses ATP and Ca2+ to control life, death and cellular signaling. Philos T R Soc B 371: 20150419–20150433. doi: 10.1098/rstb.2015.0419
|
| [153] |
154. Napoletano C, Rughetti A, Tarp MPA, et al. (2007) Tumor-associated Tn-MUC1 glycoform is internalized through the macrophage galactose-type C-type lectin and delivered to the HLA class I and II compartments in dendritic cells. Cancer Res 67: 8358–8367. doi: 10.1158/0008-5472.CAN-07-1035
|
| [154] |
155. Hiltbold EM, Vlad AM, Ciborowski P, et al. (2000) The mechanism of unresponsiveness to circulating tumor antigen MUC1 is a block in intracellular sorting and processing by dendritic cells. J Immunol 165: 3730–3741. doi: 10.4049/jimmunol.165.7.3730
|
| [155] |
156. Hanisch FG, Schwientek T, Von BergweltBaildon MS, (2003) O-Linked glycans control glycoprotein processing by antigen-presenting cells: a biochemical approach to the molecular aspects of MUC1 processing by dendritic cells. Eur J Immunol 33: 3242–3254. doi: 10.1002/eji.200324189
|
| [156] |
157. Freire T, Zhang X, Dériaud E, et al. (2010) Glycosidic Tn-based vaccines targeting dermal dendritic cells favor germinal center B-cell development and potent antibody response in the absence of adjuvant. Blood 116: 3526–3536. doi: 10.1182/blood-2010-04-279133
|
| [157] |
158. Freire T, Lo-Man R, Bay S, et al. (2011) Tn glycosylation of the MUC6 protein modulates its immunogenicity and promotes the induction of the Th17-biased T cell responses. J Biol Chem 286: 7797–7811. doi: 10.1074/jbc.M110.209742
|
| [158] |
159. Li D, Romain G, Flamar AL, et al. (2012) Targeting self- and foreign antigens to dendritic cells via DC-ASGPR generate IL-10-producing suppressive CD4+ T cells. J Exp Med 209: 109–121. doi: 10.1084/jem.20110399
|
| [159] |
160. Valladeau J, Duvert-Frances V, Pin JJ, et al. (2001) Immature human dendritic cells express asialoglycoprotein receptor isoforms for efficient receptor-mediated endocytosis. J Immunol 167: 5767–5774. doi: 10.4049/jimmunol.167.10.5767
|
| [160] | 161. Garg S, Oran A, Wajchman J, et al. (2003) Genetic tagging shows increased frequency and longevity of antigen-presenting, skin-derived dendritic cells in vivo. Nat Immunol 4: 907–912. |
| [161] | 162. Tomura M, Hata A, Matsuoka S, et al. (2014) Tracking and quantification of dendritic cell migration and antigen trafficking between the skin and lymph nodes. Sci Rep 4: 6030–6040. |
| [162] |
163. Kitano M, Yamazaki C, Takumi A, et al. (2016) Imaging of the cross-presenting dendritic cell subsets in the skin-draining lymph node. Proc Natl Acad Sci USA 113: 1044–1049. doi: 10.1073/pnas.1513607113
|
| [163] |
164. Wan YY, Flavell RA (2009) How diverse-CD4 effector T cells and their functions. J Mol Cell Biol 1: 20–36. doi: 10.1093/jmcb/mjp001
|
| [164] |
165. Lanzavecchia A (1985) Antigen-specific interaction between T and B cells. Nature 314: 537–539. doi: 10.1038/314537a0
|
| [165] |
166. Shumilina E, Huber SM, Lang F (2011) Ca2+ signaling in the regulation of dendritic cell functions. Am J Physiol Cell Ph 300: C1205–C1214. doi: 10.1152/ajpcell.00039.2011
|
| [166] |
167. Cowen DS, Lazarus HM, Shurin SB, et al. (1989) Extracellular adenosine triphosphate activates calcium mobilization in human phagocytic leukocytes and neutrophil/monocyte progenitor cells. J Clin Invest 83: 1651–. doi: 10.1172/JCI114064
|
| [167] | 168. Bretou M, Sáez PJ, Sanséau D, et al. (2017) Lysosome signaling controls the migration of dendritic cells. Sci Immunol 2: In press. |
| [168] |
169. Vukcevic M, Zorzato F, Spagnoli G, et al. (2010) Frequent calcium oscillations lead to NFAT activation in human immature dendritic cells. J Biol Chem 285: 16003–16011. doi: 10.1074/jbc.M109.066704
|
| [169] |
170. Jégouzo SAF, Quintero-Martinez, Ouyang X, et al. (2013) Organization of the extracellular portion of the macrophage galactose receptor: a trimeric cluster of simple binding sites for N-acetylgalactosamine. Glycobiology 23: 853–864. doi: 10.1093/glycob/cwt022
|
| [170] | 171. Humeau J, Bravo-San PJ, Vitale I, et al. (2017) Calcium signaling and cell cycle: progression or death. Cell Calcium 17: In press. |
| [171] |
172. Nicotera P, Orrenius S (1998) The role of calcium in apoptosis. Cell Calcium 23: 173–180. doi: 10.1016/S0143-4160(98)90116-6
|
| [172] | 173. Schwarz EC, Qu B, Hoth M (013) Calcium, cancer and killing: the role of calcium in killing cancer cells by cytotoxic T lymphocytes and natural killing cells. BBA-Mol Cell Res 1833: 1603–1611. |
| [173] |
174. Cui C, Merritt R, Fu L, et al. (2017) Targeting calcium signaling in cancer therapy. Acta Pharm Sinica B 7: 3–17. doi: 10.1016/j.apsb.2016.11.001
|
| [174] |
175. Halling DB, Liebeskind BJ, Hall AW, et al. (2016) Conserved properties of individual Ca2+-binding sites in calmodulin. Proc Nat Acad Sci USA 113: E1216–E1225. doi: 10.1073/pnas.1600385113
|
| [175] |
176. Agrawal RS, Connolly SF, Herrmann TL, et al. (2007) MHC class II levels and intracellular localization in human dendritic cells are regulated by calmodulin kinase II. J Leukocyte Biol 82: 686–699. doi: 10.1189/jlb.0107045
|
| [176] |
177. Connolly SF, Kusner DJ (2007) The regulation of dendritic cell function by calcium-signaling and its inhibition by microbial pathogens. Immunol Res 39: 115–127. doi: 10.1007/s12026-007-0076-1
|
| [177] |
178. Shi Y (2009) Serine/threonine phosphatases: mechanism through structure. Cell 139: 468–484. doi: 10.1016/j.cell.2009.10.006
|
| [178] | 179. Song MS, Salmena L, Pandolfi PP (2012) The functions and regulation of the PTEN tumor suppressor. Nat Rev Mol Cell Bio 13: 283–296. |
| [179] |
180. Cho US, Xu W (2007) Crystal structure of a protein phosphatase 2A heterotrimeric holoenzyme. Nature 445: 53–57. doi: 10.1038/nature05351
|
| [180] |
181. Feske S (2007) Calcium signaling in lymphocyte activation and disease. Nat Rev Immunol 7: 690–702. doi: 10.1038/nri2152
|
| [181] |
182. Müller MR, Rao A (2010) NFAT, immunity and cancer: a transcription factor comes of age. Nat Rev Immunol 10: 645–656. doi: 10.1038/nri2818
|
| [182] |
183. Nakanishi A, Hatano N, Fujiwara Y, et al. (2017) AMP-activated protein-kinase-mediated feedback phosphorylation controls the Ca2+/calmodulin dependence of Ca2+/CaM-dependent protein kinase kinase β. J Biol Chem 292: 19804–19813. doi: 10.1074/jbc.M117.805085
|
| [183] |
184. Gaertner TR, Kolodziej SJ, Wang D, et al. (2004) Comparative analysis of the three-dimensional structures and enzymatic properties of α, β, γ, and δ isoforms of Ca2+-calmodulin-dependent protein kinase II. J Biol Chem 279: 12484–12494. doi: 10.1074/jbc.M313597200
|
| [184] |
185. Wiede F, Dudakov JA, Lu KH, et al. (2017) PTPN2 regulates T cell lineage commitment and αβ versus γδ specification. J Exp Med 214: 2733–2758. doi: 10.1084/jem.20161903
|
| [185] |
186. Wang X, Marks CR, Perfitt TL, et al. (2017) A novel mechanism for Ca2+/calmodulin-dependent protein kinase II targeting to L-type Ca2+ channels that initiates long-range signaling to the nucleus. J Biol Chem 292: 17324–17336. doi: 10.1074/jbc.M117.788331
|
| [186] |
187. Lin MY, Zal T, Ch'En IL, et al. (2005) A pivotal role for the multifunctional calcium/calmodulin-dependent protein kinase II in T cells: from activation to unresponsiveness. J Immunol 174: 5583–5592. doi: 10.4049/jimmunol.174.9.5583
|
| [187] | 188. Ratner AJ, Bryan R, Weber A, et al. (2017) Cystic fibrosis pathogens activate Ca2+-dependent mitogen-activated protein kinase signaling pathways in airway epithelial cells. J Biol Chem 276: 19267–19275. |
| [188] | 189. Bononi A, Agnoletto C, De ME, et al. (2011) Protein kinases and phosphatases in the control of cell fate. Enz Res 2011: 329098–329113. |
| [189] |
190. Suzuki K, Hata S, Kawabata Y, et al. (2004) Structure, activation, and biology of calpain. Diabetes 53: S12–S18. doi: 10.2337/diabetes.53.2007.S12
|
| [190] | 191. Frangioni JV, Oda A, Smith M, et al. (1993) Calpain-catalyzed cleavage and subcellular relocation of protein phosphotyrosine phosphatase 1B (PTP-1B) in human platelets. EMBO J 12: 4843–4856. |
| [191] | 192. Baba Y, Kurosaki T (2016) Role of calcium signaling in B cell activation and biology. Curr Top Microbiol 393: 143–147. |
| [192] |
193. Vaeth M, Zee I, Concepcion AR, et al. (2015) Ca2+ signaling but not store-operated Ca2+ entry is required for the function of macrophages and dendritic cells. J Immunol 195: 1202–1217. doi: 10.4049/jimmunol.1403013
|
| [193] |
194. Ledderose C, Bao Y, Lidicky M, et al. (2014) Mitochondria are gate-keepers of T cell function by producing the ATP that drives purinergic signaling. J Biol Chem 289: 25936–25945. doi: 10.1074/jbc.M114.575308
|
| [194] | 195. Zizzari IG, Napoletano C, Battisti F, et al. (2015) MGL receptor and immunity: when the ligand can make the difference. J Immunol Res 2015: 450695–450702. |
| [195] |
196. van Vliet SJ, Bay S, Vuist IM, et al. (2013) MGL signaling augments TLR2-mediated responses for enhanced IL-10 and TNF-α secretion. J Leukocyte Biol 94: 315–323. doi: 10.1189/jlb.1012520
|
| [196] |
197. Nunes P, Demaurex N (2010) The role of calcium signaling in phagocytosis. J Leukocyte Biol 88: 57–68. doi: 10.1189/jlb.0110028
|
| [197] |
198. Lm VDB, Gringhuis SI, Geijtenbeek TB (2012) An evolutionary perspective on C-type lectins in infection and immunity. Ann Ny Acad Sci 1253: 149–158. doi: 10.1111/j.1749-6632.2011.06392.x
|
| [198] | 199. Kushchayev SV, Sankar T, Eggink LL, et al. (2012) Monocyte galactose/N-acetylgalactosamine-specific C-type lectin receptor stimulant immunotherapy of an experimental glioma. Part 1: stimulatory effects on blood monocytes and monocyte-derived cells of the brain. Cancer Manag Res 4: 309–323. |
| [199] | 200. Kushchayev SV, Sankar T, Eggink LL, et al. (2012) Monocyte galactose/N-acetylgalactosamine-specific C-type lectin receptor stimulant immunotherapy of an experimental glioma. Part II: combination with external radiation improves survival. Cancer Manag Res 4: 325–334. |
| [200] | 201. Roby KF, Eggink LL, Hoober JK (2017) An innovative immunotherapeutic strategy for ovarian cancer: glycomimetic peptides [abstract]. In: Proceedings of the American Association for Cancer Research Annual Meeting 2017, Washington DC. Available from: http://cancerres.aacrjournals.org/content/77/13_Supplement/170. |
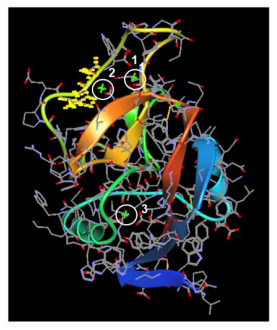

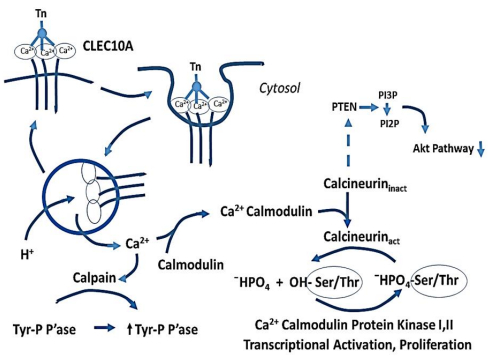
Figures(3)

Robert Cote, Laura Lynn Eggink, J. Kenneth Hoober. CLEC receptors, endocytosis and calcium signaling[J]. AIMS Allergy and Immunology, 2017, 1(4): 207-231. doi: 10.3934/Allergy.2017.4.207







 DownLoad:
DownLoad: