Citation: Seidu A. Richard, Wu Min, Zhaoliang Su, Hua-Xi Xu. Epochal neuroinflammatory role of high mobility group box 1 in central nervous system diseases[J]. AIMS Molecular Science, 2017, 4(2): 185-218. doi: 10.3934/molsci.2017.2.185

| [1] |
Fernandes-Alnemri T, Yu J-W, Datta P, et al. (2009) AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 458: 509-513. doi: 10.1038/nature07710

|
| [2] | Banjara M, Ghosh C (2017) Sterile Neuroinflammation and Strategies for Therapeutic Intervention. Int J Inflam 2017: 8385961. |
| [3] |
Lyman M, Lloyd DG, Ji X, et al. (2014) Neuroinflammation: the role and consequences. Neurosci Res 79: 1-12. doi: 10.1016/j.neures.2013.10.004
|
| [4] | Romani M, Rodman TC, Vidali G, et al. (1979) Serological analysis of species specificity in the high mobility group chromosomal proteins. J Biol Chem 254: 2918-2922. |
| [5] | Prasad S, Thakur M (1989) Distribution of high mobility group proteins in different tissues of rats during aging. Biochem Int 20: 687-695. |
| [6] |
Čabart P, Kalousek I, Jandová D, et al. (1995) Differential expression of nuclear HMG1, HMG2 proteins and H10 histone in various blood cells. Cell Biochem Funct 13: 125-133. doi: 10.1002/cbf.290130209
|
| [7] |
Seyedin SM, Pehrson JR, Cole RD (1981) Loss of chromosomal high mobility group proteins HMG1 and HMG2 when mouse neuroblastoma and Friend erythroleukemia cells become committed to differentiation. Proc Natl Acad Sci U S A 78: 5988-5992. doi: 10.1073/pnas.78.10.5988
|
| [8] |
Muller S, Ronfani L, Bianchi ME (2004) Regulated expression and subcellular localization of HMGB1, a chromatin protein with a cytokine function. J Intern Med 255: 332-343. doi: 10.1111/j.1365-2796.2003.01296.x
|
| [9] |
Kitahara T, Takeishi Y, Harada M, et al. (2008) High-mobility group box 1 restores cardiac function after myocardial infarction in transgenic mice. Cardiovasc Res 80: 40-46. doi: 10.1093/cvr/cvn163
|
| [10] |
Kang R, Zhang Q, Hou W, et al. (2014) Intracellular Hmgb1 inhibits inflammatory nucleosome release and limits acute pancreatitis in mice. Gastroenterology 146: 1097-1107. doi: 10.1053/j.gastro.2013.12.015
|
| [11] |
Huang H, Nace GW, McDonald K-A, et al. (2014) Hepatocyte specific HMGB1 deletion worsens the injury in liver ischemia/reperfusion: A role for intracellular HMGB1 in cellular protection. Hepatology (Baltimore, Md) 59: 1984. doi: 10.1002/hep.26976
|
| [12] |
Yanai H, Matsuda A, An J, et al. (2013) Conditional ablation of HMGB1 in mice reveals its protective function against endotoxemia and bacterial infection. Proc Natl Acad Sci U S A 110: 20699-20704. doi: 10.1073/pnas.1320808110
|
| [13] |
Huebener P, Gwak G-Y, Pradere J-P, et al. (2014) High-mobility group box 1 is dispensable for autophagy, mitochondrial quality control, and organ function in vivo. Cell Metab 19: 539-547. doi: 10.1016/j.cmet.2014.01.014
|
| [14] |
Chirico V, Lacquaniti A, Salpietro V, et al. (2014) High-mobility group box 1 (HMGB1) in childhood: from bench to bedside. Eur J Pediatr 173: 1123-1136. doi: 10.1007/s00431-014-2327-1
|
| [15] |
Wang K-C, Tsai C-P, Lee C-L, et al. (2012) Elevated plasma high-mobility group box 1 protein is a potential marker for neuromyelitis optica. Neuroscience 226: 510-516. doi: 10.1016/j.neuroscience.2012.08.041
|
| [16] |
Wang H, Wang K, Wang C, et al. (2013) Cerebrospinal fluid high-mobility group box protein 1 in neuromyelitis optica and multiple sclerosis. Neuroimmunomodulation 20: 113-118. doi: 10.1159/000345994
|
| [17] |
Kobori N, Clifton GL, Dash P (2002) Altered expression of novel genes in the cerebral cortex following experimental brain injury. Brain Res Mol Brain Res 104: 148-158. doi: 10.1016/S0169-328X(02)00331-5
|
| [18] |
Agnello D, Wang H, Yang H, et al. (2002) HMGB-1, a DNA-binding protein with cytokine activity, induces brain TNF and IL-6 production, and mediates anorexia and taste aversion. Cytokine 18: 231-236. doi: 10.1006/cyto.2002.0890
|
| [19] | Antoine DJ, Harris HE, Andersson U, et al. (2014) A systematic nomenclature for the redox states of high mobility group box (HMGB) proteins. Mol Med 20: 135-137. |
| [20] |
Kang R, Chen R, Zhang Q, et al. (2014) HMGB1 in health and disease. Mol Aspects Med 40: 1-116. doi: 10.1016/j.mam.2014.05.001
|
| [21] |
Andersson U, Antoine DJ, Tracey KJ (2014) The functions of HMGB1 depend on molecular localization and post-translational modifications. J Intern Med 276: 420-424. doi: 10.1111/joim.12309
|
| [22] | Su Z, Ni P, She P, et al. (2016) Bio-HMGB1 from breast cancer contributes to M-MDSC differentiation from bone marrow progenitor cells and facilitates conversion of monocytes into MDSC-like cells. Cancer Immunol Immunother 66: 391-401. |
| [23] |
Thomas JO, Stott K (2012) H1 and HMGB1: modulators of chromatin structure. Biochem Soc Trans 40: 341-346. doi: 10.1042/BST20120014
|
| [24] |
Cato L, Stott K, Watson M, et al. (2008) The interaction of HMGB1 and linker histones occurs through their acidic and basic tails. J Mol Biol 384: 1262-1272. doi: 10.1016/j.jmb.2008.10.001
|
| [25] |
Holdenrieder S, Stieber P (2009) Clinical use of circulating nucleosomes. Crit Rev Clin Lab Sci 46: 1-24. doi: 10.1080/10408360802485875
|
| [26] |
Assenberg R, Webb M, Connolly E, et al. (2008) A critical role in structure-specific DNA binding for the acetylatable lysine residues in HMGB1. Biochem J 411: 553-561. doi: 10.1042/BJ20071613
|
| [27] |
Little AJ, Corbett E, Ortega F, et al. (2013) Cooperative recruitment of HMGB1 during V (D) J recombination through interactions with RAG1 and DNA. Nucleic Acids Res 41: 3289-3301. doi: 10.1093/nar/gks1461
|
| [28] |
Das D, Scovell WM (2001) The binding interaction of HMG-1 with the TATA-binding protein/TATA complex. J Biol Chem 276: 32597-32605. doi: 10.1074/jbc.M011792200
|
| [29] |
Naghavi MH, Nowak P, Andersson J, et al. (2003) Intracellular high mobility group B1 protein (HMGB1) represses HIV-1 LTR-directed transcription in a promoter-and cell-specific manner. Virology 314: 179-189. doi: 10.1016/S0042-6822(03)00453-7
|
| [30] |
Ellwood KB, Yen Y-M, Johnson RC, et al. (2000) Mechanism for specificity by HMG-1 in enhanceosome assembly. Mol Cell Biol 20: 4359-4370. doi: 10.1128/MCB.20.12.4359-4370.2000
|
| [31] |
Mitsouras K, Wong B, Arayata C, et al. (2002) The DNA architectural protein HMGB1 displays two distinct modes of action that promote enhanceosome assembly. Mol Cell Biol 22: 4390-4401. doi: 10.1128/MCB.22.12.4390-4401.2002
|
| [32] |
Yuan F, Gu L, Guo S, et al. (2004) Evidence for involvement of HMGB1 protein in human DNA mismatch repair. J Biol Chem 279: 20935-20940. doi: 10.1074/jbc.M401931200
|
| [33] |
Robertson AB, Klungland A, Rognes T, et al. (2009) DNA repair in mammalian cells: Base excision repair: the long and short of it. Cell Mol Life Sci 66: 981-993. doi: 10.1007/s00018-009-8736-z
|
| [34] |
Prasad R, Liu Y, Deterding LJ, et al. (2007) HMGB1 is a cofactor in mammalian base excision repair. Mol Cell 27: 829-841. doi: 10.1016/j.molcel.2007.06.029
|
| [35] | Giavara S, Kosmidou E, Hande MP, et al. (2005) Yeast Nhp6A/B and mammalian Hmgb1 facilitate the maintenance of genome stability. Curr Biol 15: 68-72. |
| [36] |
Zayed H, Izsvák Z, Khare D, et al. (2003) The DNA‐bending protein HMGB1 is a cellular cofactor of Sleeping Beauty transposition. Nucleic Acids Res 31: 2313-2322. doi: 10.1093/nar/gkg341
|
| [37] |
de Silva S, Lotta LT, Jr., Burris CA, et al. (2010) Virion-associated cofactor high-mobility group DNA-binding protein-1 facilitates transposition from the herpes simplex virus/Sleeping Beauty amplicon vector platform. Hum Gene Ther 21: 1615-1622. doi: 10.1089/hum.2010.022
|
| [38] |
Shen Y, Peng H, Pan S, et al. (2010) Interaction of DNA/nuclear protein/polycation and the terplexes for gene delivery. Nanotechnology 21: 045102. doi: 10.1088/0957-4484/21/4/045102
|
| [39] |
Siu YS, Li L, Leung MF, et al. (2012) Polyethylenimine-based amphiphilic core-shell nanoparticles: study of gene delivery and intracellular trafficking. Biointerphases 7: 16. doi: 10.1007/s13758-011-0016-4
|
| [40] |
Yi WJ, Yang J, Li C, et al. (2012) Enhanced nuclear import and transfection efficiency of TAT peptide-based gene delivery systems modified by additional nuclear localization signals. Bioconjug Chem 23: 125-134. doi: 10.1021/bc2005472
|
| [41] |
Weber MD, Frank MG, Tracey KJ, et al. (2015) Stress induces the danger-associated molecular pattern HMGB-1 in the hippocampus of male Sprague Dawley rats: a priming stimulus of microglia and the NLRP3 inflammasome. J Neurosci 35: 316-324. doi: 10.1523/JNEUROSCI.3561-14.2015
|
| [42] |
Tang D, Kang R, Livesey KM, et al. (2010) Endogenous HMGB1 regulates autophagy. J Cell Biol 190: 881-892. doi: 10.1083/jcb.200911078
|
| [43] |
Lee H, Shin N, Song M, et al. (2010) Analysis of nuclear high mobility group box 1 (HMGB1)-binding proteins in colon cancer cells: clustering with proteins involved in secretion and extranuclear function. J Proteome Res 9: 4661-4670. doi: 10.1021/pr100386r
|
| [44] |
Dintilhac A, Bernues J (2002) HMGB1 interacts with many apparently unrelated proteins by recognizing short amino acid sequences. J Biol Chem 277: 7021-7028. doi: 10.1074/jbc.M108417200
|
| [45] |
Kang R, Livesey KM, Zeh HJ, 3rd, et al. (2011) Metabolic regulation by HMGB1-mediated autophagy and mitophagy. Autophagy 7: 1256-1258. doi: 10.4161/auto.7.10.16753
|
| [46] | Tang D, Billiar TR, Lotze MT (2012) A Janus tale of two active high mobility group box 1 (HMGB1) redox states. Mol Med 18: 1360-1362. |
| [47] |
Fang WH, Yao YM, Shi ZG, et al. (2002) The significance of changes in high mobility group-1 protein mRNA expression in rats after thermal injury. Shock 17: 329-333. doi: 10.1097/00024382-200204000-00016
|
| [48] |
Wang H, Bloom O, Zhang M, et al. (1999) HMG-1 as a late mediator of endotoxin lethality in mice. Science 285: 248-251. doi: 10.1126/science.285.5425.248
|
| [49] |
Wang H, Vishnubhakat JM, Bloom O, et al. (1999) Proinflammatory cytokines (tumor necrosis factor and interleukin 1) stimulate release of high mobility group protein-1 by pituicytes. Surgery 126: 389-392. doi: 10.1016/S0039-6060(99)70182-0
|
| [50] |
Andersson U, Wang H, Palmblad K, et al. (2000) High mobility group 1 protein (HMG-1) stimulates proinflammatory cytokine synthesis in human monocytes. J Exp Med 192: 565-570. doi: 10.1084/jem.192.4.565
|
| [51] |
Yang H, Wang H, Tracey KJ (2001) HMG-1 rediscovered as a cytokine. Shock 15: 247-253. doi: 10.1097/00024382-200115040-00001
|
| [52] | Alexander HR, Doherty GM, Venzon DJ, et al. (1992) Recombinant interleukin-1 receptor antagonist (IL-1ra): effective therapy against gram-negative sepsis in rats. Surgery 112: 188-193; discussion 193-184. |
| [53] |
Libert C, Vink A, Coulie P, et al. (1992) Limited involvement of interleukin‐6 in the pathogenesis of lethal septic shock as revealed by the effect of monoclonal antibodies against interleukin‐6 or its receptor in various murine models. Eur J Immunol 22: 2625-2630. doi: 10.1002/eji.1830221023
|
| [54] |
Abraham E, Arcaroli J, Carmody A, et al. (2000) Cutting edge: HMG-1 as a mediator of acute lung inflammation. J Immunol 165: 2950-2954. doi: 10.4049/jimmunol.165.6.2950
|
| [55] |
Wagner R, Myers RR (1996) Endoneurial injection of TNF-alpha produces neuropathic pain behaviors. Neuroreport 7: 2897-2901. doi: 10.1097/00001756-199611250-00018
|
| [56] |
Eliav E, Herzberg U, Ruda MA, et al. (1999) Neuropathic pain from an experimental neuritis of the rat sciatic nerve. Pain 83: 169-182. doi: 10.1016/S0304-3959(99)00102-5
|
| [57] |
Chacur M, Milligan ED, Gazda LS, et al. (2001) A new model of sciatic inflammatory neuritis (SIN): induction of unilateral and bilateral mechanical allodynia following acute unilateral peri-sciatic immune activation in rats. Pain 94: 231-244. doi: 10.1016/S0304-3959(01)00354-2
|
| [58] |
Shimommura Y, Shimizu H, Takahashi M, et al. (1990) Effects of peripheral administration of recombinant human interleukin-1 beta on feeding behavior of the rat. Life Sci 47: 2185-2192. doi: 10.1016/0024-3205(90)90148-K
|
| [59] |
Crestani F, Seguy F, Dantzer R (1991) Behavioural effects of peripherally injected interleukin-1: role of prostaglandins. Brain Res 542: 330-335. doi: 10.1016/0006-8993(91)91587-Q
|
| [60] |
Dunn AJ, Antoon M, Chapman Y (1991) Reduction of exploratory behavior by intraperitoneal injection of interleukin-1 involves brain corticotropin-releasing factor. Brain Res Bull 26: 539-542. doi: 10.1016/0361-9230(91)90092-X
|
| [61] | Kluger MJ (1991) Fever: role of pyrogens and cryogens. Physiol Rev 71: 93-127. |
| [62] |
Oka T, Aou S, Hori T (1993) Intracerebroventricular injection of interleukin-1β induces hyperalgesia in rats. Brain Res 624: 61-68. doi: 10.1016/0006-8993(93)90060-Z
|
| [63] |
Bluthe R-M, Michaud B, Kelley KW, et al. (1996) Vagotomy attenuates behavioural effects of interleukin-1 injected peripherally but not centrally. Neuroreport 7: 1485-1488. doi: 10.1097/00001756-199606170-00008
|
| [64] |
Reeve AJ, Patel S, Fox A, et al. (2000) Intrathecally administered endotoxin or cytokines produce allodynia, hyperalgesia and changes in spinal cord neuronal responses to nociceptive stimuli in the rat. Eur J Pain 4: 247-257. doi: 10.1053/eujp.2000.0177
|
| [65] |
Yirmiya R (1996) Endotoxin produces a depressive-like episode in rats. Brain Res 711: 163-174. doi: 10.1016/0006-8993(95)01415-2
|
| [66] |
Coelho A, Fioramonti J, Bueno L (2000) Brain interleukin-1beta and tumor necrosis factor-alpha are involved in lipopolysaccharide-induced delayed rectal allodynia in awake rats. Brain Res Bull 52: 223-228. doi: 10.1016/S0361-9230(00)00269-0
|
| [67] |
Turrin NP, Gayle D, Ilyin SE, et al. (2001) Pro-inflammatory and anti-inflammatory cytokine mRNA induction in the periphery and brain following intraperitoneal administration of bacterial lipopolysaccharide. Brain Res Bull 54: 443-453. doi: 10.1016/S0361-9230(01)00445-2
|
| [68] |
Gabellec M-M, Griffais R, Fillion G, et al. (1995) Expression of interleukin 1α, interleukin 1β and interleukin 1 receptor antagonist mRNA in mouse brain: regulation by bacterial lipopolysaccharide (LPS) treatment. Mol Brain Res 31: 122-130. doi: 10.1016/0169-328X(95)00042-Q
|
| [69] | Laye S, Gheusi G, Cremona S, et al. (2000) Endogenous brain IL-1 mediates LPS-induced anorexia and hypothalamic cytokine expression. Am J Physiol Regul Integr Comp Physiol 279: R93-98. |
| [70] |
Bluthe RM, Dantzer R, Kelley KW (1997) Central mediation of the effects of interleukin-1 on social exploration and body weight in mice. Psychoneuroendocrinology 22: 1-11. doi: 10.1016/S0306-4530(96)00042-X
|
| [71] |
Rothwell NJ (1988) Central effects of TNFα on thermogenesis and fever in the rat. Biosci Rep 8: 345-352. doi: 10.1007/BF01115225
|
| [72] | Dascombe MJ, Rothwell NJ, Sagay BO, et al. (1989) Pyrogenic and thermogenic effects of interleukin 1 beta in the rat. Am J Physiol 256: E7-11. |
| [73] |
Walter JS, Meyers P, Krueger JM (1989) Microinjection of interleukin-1 into brain: separation of sleep and fever responses. Physiol Behav 45: 169-176. doi: 10.1016/0031-9384(89)90181-9
|
| [74] |
O'Connor KA, Hansen MK, Pugh CR, et al. (2003) Further characterization of high mobility group box 1 (HMGB1) as a proinflammatory cytokine: central nervous system effects. Cytokine 24: 254-265. doi: 10.1016/j.cyto.2003.08.001
|
| [75] | Fiebich BL, Schleicher S, Spleiss O, et al. (2001) Mechanisms of prostaglandin E2-induced interleukin-6 release in astrocytes: possible involvement of EP4-like receptors, p38 mitogen-activated protein kinase and protein kinase C. J Neurochem 79: 950-958. |
| [76] |
Newton R, Kuitert LM, Bergmann M, et al. (1997) Evidence for involvement of NF-kappaB in the transcriptional control of COX-2 gene expression by IL-1beta. Biochem Biophys Res Commun 237: 28-32. doi: 10.1006/bbrc.1997.7064
|
| [77] |
Hori O, Brett J, Slattery T, et al. (1995) The receptor for advanced glycation end products (RAGE) is a cellular binding site for amphoterin mediation of neurite outgrowth and co-expression of rage and amphoterin in the developing nervous system. J Biol Chem 270: 25752-25761. doi: 10.1074/jbc.270.43.25752
|
| [78] |
Fiuza C, Bustin M, Talwar S, et al. (2003) Inflammation-promoting activity of HMGB1 on human microvascular endothelial cells. Blood 101: 2652-2660. doi: 10.1182/blood-2002-05-1300
|
| [79] |
Park JS, Arcaroli J, Yum HK, et al. (2003) Activation of gene expression in human neutrophils by high mobility group box 1 protein. Am J Physiol Cell Physiol 284: C870-879. doi: 10.1152/ajpcell.00322.2002
|
| [80] |
Chen GY, Tang J, Zheng P, et al. (2009) CD24 and Siglec-10 selectively repress tissue damage-induced immune responses. Science 323: 1722-1725. doi: 10.1126/science.1168988
|
| [81] |
Tian J, Avalos AM, Mao S-Y, et al. (2007) Toll-like receptor 9–dependent activation by DNA-containing immune complexes is mediated by HMGB1 and RAGE. Nat Immunol 8: 487-496. doi: 10.1038/ni1457
|
| [82] |
Gao H-M, Zhou H, Zhang F, et al. (2011) HMGB1 acts on microglia Mac1 to mediate chronic neuroinflammation that drives progressive neurodegeneration. J Neurosci 31: 1081-1092. doi: 10.1523/JNEUROSCI.3732-10.2011
|
| [83] |
Fu GX, Chen AF, Zhong Y, et al. (2016) Decreased serum level of HMGB1 and MyD88 during human aging progress in healthy individuals. Aging Clin Exp Res 28: 175-180. doi: 10.1007/s40520-015-0402-8
|
| [84] |
Tang AH, Brunn GJ, Cascalho M, et al. (2007) Pivotal advance: endogenous pathway to SIRS, sepsis, and related conditions. J Leukoc Biol 82: 282-285. doi: 10.1189/jlb.1206752
|
| [85] | Park JS, Gamboni-Robertson F, He Q, et al. (2006) High mobility group box 1 protein interacts with multiple Toll-like receptors. Am J Physiol Cell Physiol 290: C917-924. |
| [86] |
Imai Y, Kuba K, Neely GG, et al. (2008) Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 133: 235-249. doi: 10.1016/j.cell.2008.02.043
|
| [87] |
Salminen A, Ojala J, Kaarniranta K, et al. (2012) Mitochondrial dysfunction and oxidative stress activate inflammasomes: impact on the aging process and age-related diseases. Cell Mol Life Sci 69: 2999-3013. doi: 10.1007/s00018-012-0962-0
|
| [88] |
Morinaga Y, Yanagihara K, Nakamura S, et al. (2010) Legionella pneumophila induces cathepsin B-dependent necrotic cell death with releasing high mobility group box1 in macrophages. Respir Res 11: 158. doi: 10.1186/1465-9921-11-158
|
| [89] | Rauvala H, Pihlaskari R (1987) Isolation and some characteristics of an adhesive factor of brain that enhances neurite outgrowth in central neurons. J Biol Chem 262: 16625-16635. |
| [90] |
Guazzi S, Strangio A, Franzi AT, et al. (2003) HMGB1, an architectural chromatin protein and extracellular signalling factor, has a spatially and temporally restricted expression pattern in mouse brain. Gene Expr Patterns 3: 29-33. doi: 10.1016/S1567-133X(02)00093-5
|
| [91] |
Enokido Y, Yoshitake A, Ito H, et al. (2008) Age-dependent change of HMGB1 and DNA double-strand break accumulation in mouse brain. Biochem Biophys Res Commun 376: 128-133. doi: 10.1016/j.bbrc.2008.08.108
|
| [92] |
Fang P, Schachner M, Shen YQ (2012) HMGB1 in development and diseases of the central nervous system. Mol Neurobiol 45: 499-506. doi: 10.1007/s12035-012-8264-y
|
| [93] |
Fonken LK, Frank MG, Kitt MM, et al. (2016) The Alarmin HMGB1 Mediates Age-Induced Neuroinflammatory Priming. J Neurosci 36: 7946-7956. doi: 10.1523/JNEUROSCI.1161-16.2016
|
| [94] |
O'Donovan MC (1993) A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. Cell 72: 971-983. doi: 10.1016/0092-8674(93)90585-E
|
| [95] |
Hodges A, Strand AD, Aragaki AK, et al. (2006) Regional and cellular gene expression changes in human Huntington's disease brain. Hum Mol Genet 15: 965-977. doi: 10.1093/hmg/ddl013
|
| [96] |
Crotti A, Benner C, Kerman BE, et al. (2014) Mutant Huntingtin promotes autonomous microglia activation via myeloid lineage-determining factors. Nat Neurosci 17: 513-521. doi: 10.1038/nn.3668
|
| [97] |
Crotti A, Glass CK (2015) The choreography of neuroinflammation in Huntington's disease. Trends Immunol 36: 364-373. doi: 10.1016/j.it.2015.04.007
|
| [98] |
Min HJ, Ko EA, Wu J, et al. (2013) Chaperone-like activity of high-mobility group box 1 protein and its role in reducing the formation of polyglutamine aggregates. J Immunol 190: 1797-1806. doi: 10.4049/jimmunol.1202472
|
| [99] |
Qi M-L, Tagawa K, Enokido Y, et al. (2007) Proteome analysis of soluble nuclear proteins reveals that HMGB1/2 suppress genotoxic stress in polyglutamine diseases. Nat Cell Biol 9: 402-414. doi: 10.1038/ncb1553
|
| [100] |
Goula AV, Berquist BR, Wilson DM, 3rd, et al. (2009) Stoichiometry of base excision repair proteins correlates with increased somatic CAG instability in striatum over cerebellum in Huntington's disease transgenic mice. PLoS Genet 5: e1000749. doi: 10.1371/journal.pgen.1000749
|
| [101] |
Liu Y, Prasad R, Beard WA, et al. (2009) Coordination between polymerase β and FEN1 can modulate CAG repeat expansion. J Biol Chem 284: 28352-28366. doi: 10.1074/jbc.M109.050286
|
| [102] |
McAlpine FE, Lee JK, Harms AS, et al. (2009) Inhibition of soluble TNF signaling in a mouse model of Alzheimer's disease prevents pre-plaque amyloid-associated neuropathology. Neurobiol Dis 34: 163-177. doi: 10.1016/j.nbd.2009.01.006
|
| [103] |
Frank-Cannon TC, Alto LT, McAlpine FE, et al. (2009) Does neuroinflammation fan the flame in neurodegenerative diseases? Mol Neurodegener 4: 47. doi: 10.1186/1750-1326-4-47
|
| [104] | Akiyama H, Arai T, Kondo H, et al. (2000) Cell mediators of inflammation in the Alzheimer disease brain. Alzheimer Dis Assoc Disord 14 Suppl 1: S47-53. |
| [105] | Buchanan MM, Hutchinson M, Watkins LR, et al. (2010) Toll-like receptor 4 in CNS pathologies. J Neurochem 114: 13-27. |
| [106] |
Du Yan S, Chen X, Fu J, et al. (1996) RAGE and amyloid-β peptide neurotoxicity in Alzheimer's disease. Nature 382: 685-691. doi: 10.1038/382685a0
|
| [107] |
Mazarati A, Maroso M, Iori V, et al. (2011) High-mobility group box-1 impairs memory in mice through both toll-like receptor 4 and receptor for advanced glycation end products. Exp Neurol 232: 143-148. doi: 10.1016/j.expneurol.2011.08.012
|
| [108] |
Takata K, Kitamura Y, Kakimura J, et al. (2003) Role of high mobility group protein-1 (HMG1) in amyloid-beta homeostasis. Biochem Biophys Res Commun 301: 699-703. doi: 10.1016/S0006-291X(03)00024-X
|
| [109] | Takata K, Takada T, Ito A, et al. (2012) Microglial Amyloid-beta1-40 Phagocytosis Dysfunction Is Caused by High-Mobility Group Box Protein-1: Implications for the Pathological Progression of Alzheimer's Disease. Int J Alzheimers Dis 2012: 685739. |
| [110] | Richard SA, Wu M, Lin D (2014) Traumatic Subdural Effusion Evolving into Chronic Subdural Hematoma. Open J Mod Neurosurg 5: 12. |
| [111] | Woodcock T, Morganti-Kossmann MC (2013) The role of markers of inflammation in traumatic brain injury. Front Neurol 4: 18. |
| [112] |
Laird MD, Shields JS, Sukumari-Ramesh S, et al. (2014) High mobility group box protein-1 promotes cerebral edema after traumatic brain injury via activation of toll-like receptor 4. Glia 62: 26-38. doi: 10.1002/glia.22581
|
| [113] |
Richard SA, Min W, Su Z, et al. (2017) High Mobility Group Box 1 and Traumatic Brain Injury. J Behav Brain Sci 7: 50. doi: 10.4236/jbbs.2017.72006
|
| [114] | Su X, Wang H, Zhao J, et al. (2011) Beneficial effects of ethyl pyruvate through inhibiting high-mobility group box 1 expression and TLR4/NF-B pathway after traumatic brain injury in the rat. Mediators Inflamm 2011. |
| [115] |
Wang K-Y, Yu G-F, Zhang Z-Y, et al. (2012) Plasma high-mobility group box 1 levels and prediction of outcome in patients with traumatic brain injury. Clin Chim Acta 413: 1737-1741. doi: 10.1016/j.cca.2012.07.002
|
| [116] |
Au AK, Aneja RK, Bell MJ, et al. (2012) Cerebrospinal fluid levels of high-mobility group box 1 and cytochrome C predict outcome after pediatric traumatic brain injury. J Neurotrauma 29: 2013-2021. doi: 10.1089/neu.2011.2171
|
| [117] |
Carty M, Bowie AG (2011) Evaluating the role of Toll-like receptors in diseases of the central nervous system. Biochem Pharmacol 81: 825-837. doi: 10.1016/j.bcp.2011.01.003
|
| [118] |
Ding Q, Keller JN (2005) Evaluation of rage isoforms, ligands, and signaling in the brain. Biochim Biophys Acta 1746: 18-27. doi: 10.1016/j.bbamcr.2005.08.006
|
| [119] |
Okuma Y, Liu K, Wake H, et al. (2012) Anti–high mobility group box‐1 antibody therapy for traumatic brain injury. Ann Neurol 72: 373-384. doi: 10.1002/ana.23602
|
| [120] |
Litvan I, Halliday G, Hallett M, et al. (2007) The etiopathogenesis of Parkinson disease and suggestions for future research. Part I. J Neuropathol Exp Neurol 66: 251-257. doi: 10.1097/nen.0b013e3180415e42
|
| [121] |
Gerhard A, Pavese N, Hotton G, et al. (2006) In vivo imaging of microglial activation with [11 C](R)-PK11195 PET in idiopathic Parkinson's disease. Neurobiol Dis 21: 404-412. doi: 10.1016/j.nbd.2005.08.002
|
| [122] | Lindersson EK, Hojrup P, Gai WP, et al. (2004) alpha-Synuclein filaments bind the transcriptional regulator HMGB-1. Neuroreport 15: 2735-2739. |
| [123] |
Song J-X, Lu J-H, Liu L-F, et al. (2014) HMGB1 is involved in autophagy inhibition caused by SNCA/α-synuclein overexpression: a process modulated by the natural autophagy inducer corynoxine B. Autophagy 10: 144-154. doi: 10.4161/auto.26751
|
| [124] |
Zhang J, Niu N, Wang M, et al. (2013) Neuron-derived IgG protects dopaminergic neurons from insult by 6-OHDA and activates microglia through the FcγR I and TLR4 pathways. Int J Biochem Cell Biol 45: 1911-1920. doi: 10.1016/j.biocel.2013.06.005
|
| [125] |
Santoro M, Maetzler W, Stathakos P, et al. (2016) In-vivo evidence that high mobility group box 1 exerts deleterious effects in the 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine model and Parkinson's disease which can be attenuated by glycyrrhizin. Neurobiol Dis 91: 59-68. doi: 10.1016/j.nbd.2016.02.018
|
| [126] | Sasaki T, Liu K, Agari T, et al. (2016) Anti-high mobility group box 1 antibody exerts neuroprotection in a rat model of Parkinson's disease. Exp Neurol 275 Pt 1: 220-231. |
| [127] |
Furuya N, Yu J, Byfield M, et al. (2005) The evolutionarily conserved domain of Beclin 1 is required for Vps34 binding, autophagy, and tumor suppressor function. Autophagy 1: 46-52. doi: 10.4161/auto.1.1.1542
|
| [128] |
Funderburk SF, Wang QJ, Yue Z (2010) The Beclin 1–VPS34 complex–at the crossroads of autophagy and beyond. Trends Cell Biol 20: 355-362. doi: 10.1016/j.tcb.2010.03.002
|
| [129] |
Kang R, Livesey KM, Zeh HJ, et al. (2010) HMGB1: a novel Beclin 1-binding protein active in autophagy. Autophagy 6: 1209-1211. doi: 10.4161/auto.6.8.13651
|
| [130] | Huang J, Yang J, Shen Y, et al. (2017) HMGB1 Mediates Autophagy Dysfunction via Perturbing Beclin1-Vps34 Complex in Dopaminergic Cell Model. Front Mol Neurosci 10: 13. |
| [131] |
Constantinescu CS, Farooqi N, O'brien K, et al. (2011) Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Brit J Pharmacol 164: 1079-1106. doi: 10.1111/j.1476-5381.2011.01302.x
|
| [132] |
McFarland HF, Martin R (2007) Multiple sclerosis: a complicated picture of autoimmunity. Nat Immunol 8: 913-919. doi: 10.1038/ni1507
|
| [133] |
Andersson Å, Covacu R, Sunnemark D, et al. (2008) Pivotal advance: HMGB1 expression in active lesions of human and experimental multiple sclerosis. J Leukoc Biol 84: 1248-1255. doi: 10.1189/jlb.1207844
|
| [134] |
Sternberg Z, Sternberg D, Chichelli T, et al. (2016) High-mobility group box 1 in multiple sclerosis. Immunol Res 64: 385-391. doi: 10.1007/s12026-015-8673-x
|
| [135] | Leech S, Kirk J, Plumb J, et al. (2007) Persistent endothelial abnormalities and blood-brain barrier leak in primary and secondary progressive multiple sclerosis. Neuropathol Appl Neurobiol 33: 86-98. |
| [136] |
Šestan N (2012) The emerging biology of autism spectrum disorders. Science 337: 1301-1303. doi: 10.1126/science.1224989
|
| [137] | Madore C, Leyrolle Q, Lacabanne C, et al. (2016) Neuroinflammation in Autism: Plausible Role of Maternal Inflammation, Dietary Omega 3, and Microbiota. Neural Plast 2016: 3597209. |
| [138] |
Voineagu I, Wang X, Johnston P, et al. (2011) Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature 474: 380-384. doi: 10.1038/nature10110
|
| [139] |
Odell D, Maciulis A, Cutler A, et al. (2005) Confirmation of the association of the C4B null allelle in autism. Hum Immunol 66: 140-145. doi: 10.1016/j.humimm.2004.11.002
|
| [140] |
Oddi D, Crusio WE, D'Amato FR, et al. (2013) Monogenic mouse models of social dysfunction: implications for autism. Behav Brain Res 251: 75-84. doi: 10.1016/j.bbr.2013.01.002
|
| [141] | Shcheglovitov A, Shcheglovitova O, Yazawa M, et al. (2013) SHANK3 and IGF1 restore synaptic deficits in neurons from 22q13 deletion syndrome patients. Nature 503: 267-271. |
| [142] |
Emanuele E, Boso M, Brondino N, et al. (2010) Increased serum levels of high mobility group box 1 protein in patients with autistic disorder. Prog Neuropsychopharmacol Biol Psychiatry 34: 681-683. doi: 10.1016/j.pnpbp.2010.03.020
|
| [143] | Junaid MA, Kowal D, Barua M, et al. (2004) Proteomic studies identified a single nucleotide polymorphism in glyoxalase I as autism susceptibility factor. Am J Med Genet A 131: 11-17. |
| [144] |
Enstrom AM, Onore CE, Van de Water JA, et al. (2010) Differential monocyte responses to TLR ligands in children with autism spectrum disorders. Brain Behav Immun 24: 64-71. doi: 10.1016/j.bbi.2009.08.001
|
| [145] |
Barua M, Jenkins EC, Chen W, et al. (2011) Glyoxalase I polymorphism rs2736654 causing the Ala111Glu substitution modulates enzyme activity--implications for autism. Autism Res 4: 262-270. doi: 10.1002/aur.197
|
| [146] |
Henkel JS, Engelhardt JI, Siklos L, et al. (2004) Presence of dendritic cells, MCP-1, and activated microglia/macrophages in amyotrophic lateral sclerosis spinal cord tissue. Ann Neurol 55: 221-235. doi: 10.1002/ana.10805
|
| [147] |
Weydt P, Yuen EC, Ransom BR, et al. (2004) Increased cytotoxic potential of microglia from ALS-transgenic mice. Glia 48: 179-182. doi: 10.1002/glia.20062
|
| [148] |
Casula M, Iyer AM, Spliet WG, et al. (2011) Toll-like receptor signaling in amyotrophic lateral sclerosis spinal cord tissue. Neuroscience 179: 233-243. doi: 10.1016/j.neuroscience.2011.02.001
|
| [149] |
Lo Coco D, Veglianese P, Allievi E, et al. (2007) Distribution and cellular localization of high mobility group box protein 1 (HMGB1) in the spinal cord of a transgenic mouse model of ALS. Neurosci Lett 412: 73-77. doi: 10.1016/j.neulet.2006.10.063
|
| [150] |
Hwang C-S, Liu G-T, Chang MD-T, et al. (2013) Elevated serum autoantibody against high mobility group box 1 as a potent surrogate biomarker for amyotrophic lateral sclerosis. Neurobiol Dis 58: 13-18. doi: 10.1016/j.nbd.2013.04.013
|
| [151] | Costa GMF, Leite CMdA (2015) Trigeminal neuralgia: peripheral and central mechanisms. Revista Dor 16: 297-301. |
| [152] |
Julius D, Basbaum AI (2001) Molecular mechanisms of nociception. Nature 413: 203-210. doi: 10.1038/35093019
|
| [153] |
Austin PJ, Moalem-Taylor G (2010) The neuro-immune balance in neuropathic pain: involvement of inflammatory immune cells, immune-like glial cells and cytokines. J Neuroimmunol 229: 26-50. doi: 10.1016/j.jneuroim.2010.08.013
|
| [154] |
Karatas H, Erdener SE, Gursoy-Ozdemir Y, et al. (2013) Spreading depression triggers headache by activating neuronal Panx1 channels. Science 339: 1092-1095. doi: 10.1126/science.1231897
|
| [155] | Kunkler PE, Hulse RE, Kraig RP (2004) Multiplexed cytokine protein expression profiles from spreading depression in hippocampal organotypic cultures. J Cereb Blood Flow Metab 24: 829-839. |
| [156] | Jander S, Schroeter M, Peters O, et al. (2001) Cortical spreading depression induces proinflammatory cytokine gene expression in the rat brain. J Cereb Blood Flow Metab 21: 218-225. |
| [157] |
Lana-Peixoto MA (2008) Devic's neuromyelitis optica: a critical review. Arq Neuropsiquiatr 66: 120-138. doi: 10.1590/S0004-282X2008000100034
|
| [158] |
Correale J, Fiol M (2004) Activation of humoral immunity and eosinophils in neuromyelitis optica. Neurology 63: 2363-2370. doi: 10.1212/01.WNL.0000148481.80152.BF
|
| [159] |
Ishizu T, Osoegawa M, Mei FJ, et al. (2005) Intrathecal activation of the IL-17/IL-8 axis in opticospinal multiple sclerosis. Brain 128: 988-1002. doi: 10.1093/brain/awh453
|
| [160] |
Uzawa A, Mori M, Masuda S, et al. (2013) CSF high-mobility group box 1 is associated with intrathecal inflammation and astrocytic damage in neuromyelitis optica. J Neurol Neurosurg Psychiatry 84: 517-522. doi: 10.1136/jnnp-2012-304039
|
| [161] |
Solmi M, Veronese N, Favaro A, et al. (2015) Inflammatory cytokines and anorexia nervosa: A meta-analysis of cross-sectional and longitudinal studies. Psychoneuroendocrinology 51: 237-252. doi: 10.1016/j.psyneuen.2014.09.031
|
| [162] |
Sadagurski M, Norquay L, Farhang J, et al. (2010) Human IL6 enhances leptin action in mice. Diabetologia 53: 525-535. doi: 10.1007/s00125-009-1580-8
|
| [163] |
Senaris RM, Trujillo ML, Navia B, et al. (2011) Interleukin-6 regulates the expression of hypothalamic neuropeptides involved in body weight in a gender-dependent way. J Neuroendocrinol 23: 675-686. doi: 10.1111/j.1365-2826.2011.02158.x
|
| [164] |
Inui A (2001) Eating behavior in anorexia nervosa--an excess of both orexigenic and anorexigenic signalling? Mol Psychiatry 6: 620. doi: 10.1038/sj.mp.4000944
|
| [165] |
Scheede-Bergdahl C, Watt HL, Trutschnigg B, et al. (2012) Is IL-6 the best pro-inflammatory biomarker of clinical outcomes of cancer cachexia? Clin Nutr 31: 85-88. doi: 10.1016/j.clnu.2011.07.010
|
| [166] |
Solmi M, Santonastaso P, Caccaro R, et al. (2013) A case of anorexia nervosa with comorbid Crohn's disease: beneficial effects of anti-TNF-alpha therapy? Int J Eat Disord 46: 639-641. doi: 10.1002/eat.22153
|
| [167] |
Hargrave SL, Kinzig KP (2012) Repeated gastric distension alters food intake and neuroendocrine profiles in rats. Physiol Behav 105: 975-981. doi: 10.1016/j.physbeh.2011.11.006
|
| [168] | Sedlackova D, Kopeckova J, Papezova H, et al. (2011) Changes of plasma obestatin, ghrelin and NPY in anorexia and bulimia nervosa patients before and after a high-carbohydrate breakfast. Physiol Res 60: 165. |
| [169] |
Cuntz U, Enck P, Frühauf E, et al. (2013) Cholecystokinin revisited: CCK and the hunger trap in anorexia nervosa. PloS One 8: e54457. doi: 10.1371/journal.pone.0054457
|
| [170] | Terra X, Auguet T, Aguera Z, et al. (2013) Adipocytokine levels in women with anorexia nervosa. Relationship with weight restoration and disease duration. Int J Eat Disord 46: 855-861. |
| [171] |
Hughes EK, Goldschmidt AB, Labuschagne Z, et al. (2013) Eating disorders with and without comorbid depression and anxiety: similarities and differences in a clinical sample of children and adolescents. Eur Eat Disord Rev 21: 386-394. doi: 10.1002/erv.2234
|
| [172] |
Yasuhara D, Hashiguchi T, Kawahara K, et al. (2007) High mobility group box 1 and refeeding-resistance in anorexia nervosa. Mol Psychiatry 12: 976-977. doi: 10.1038/sj.mp.4002050
|
| [173] |
Vezzani A (2014) Epilepsy and inflammation in the brain: overview and pathophysiology. Epilepsy Curr 14: 3-7. doi: 10.5698/1535-7511-14.s2.3
|
| [174] |
Gorter JA, van Vliet EA, Aronica E, et al. (2006) Potential new antiepileptogenic targets indicated by microarray analysis in a rat model for temporal lobe epilepsy. J Neurosci 26: 11083-11110. doi: 10.1523/JNEUROSCI.2766-06.2006
|
| [175] |
Ravizza T, Vezzani A (2006) Status epilepticus induces time-dependent neuronal and astrocytic expression of interleukin-1 receptor type I in the rat limbic system. Neuroscience 137: 301-308. doi: 10.1016/j.neuroscience.2005.07.063
|
| [176] |
Turrin NP, Rivest S (2004) Innate immune reaction in response to seizures: implications for the neuropathology associated with epilepsy. Neurobiol Dis 16: 321-334. doi: 10.1016/j.nbd.2004.03.010
|
| [177] |
Vezzani A, French J, Bartfai T, et al. (2011) The role of inflammation in epilepsy. Nat Rev Neurol 7: 31-40. doi: 10.1038/nrneurol.2010.178
|
| [178] |
Maroso M, Balosso S, Ravizza T, et al. (2010) Toll-like receptor 4 and high-mobility group box-1 are involved in ictogenesis and can be targeted to reduce seizures. Nat Med 16: 413-419. doi: 10.1038/nm.2127
|
| [179] |
Liang Y, Lei Z, Zhang H, et al. (2014) Toll-like receptor 4 is associated with seizures following ischemia with hyperglycemia. Brain Res 1590: 75-84. doi: 10.1016/j.brainres.2014.09.020
|
| [180] |
Allan SM, Tyrrell PJ, Rothwell NJ (2005) Interleukin-1 and neuronal injury. Nat Rev Immunol 5: 629-640. doi: 10.1038/nri1664
|
| [181] |
Van Vliet E, da Costa Araujo S, Redeker S, et al. (2007) Blood–brain barrier leakage may lead to progression of temporal lobe epilepsy. Brain 130: 521-534. doi: 10.1093/brain/awl318
|
| [182] |
Oby E, Janigro D (2006) The blood–brain barrier and epilepsy. Epilepsia 47: 1761-1774. doi: 10.1111/j.1528-1167.2006.00817.x
|
| [183] |
Choi J, Min HJ, Shin JS (2011) Increased levels of HMGB1 and pro-inflammatory cytokines in children with febrile seizures. J Neuroinflammation 8: 135. doi: 10.1186/1742-2094-8-135
|
| [184] |
Kleen JK, Holmes GL (2010) Taming TLR4 may ease seizures. Nat Med 16: 369-370. doi: 10.1038/nm0410-369
|
| [185] |
Balosso S, Liu J, Bianchi ME, et al. (2014) Disulfide-containing high mobility group box-1 promotes N-methyl-D-aspartate receptor function and excitotoxicity by activating Toll-like receptor 4-dependent signaling in hippocampal neurons. Antioxid Redox Signal 21: 1726-1740. doi: 10.1089/ars.2013.5349
|
| [186] |
Iori V, Maroso M, Rizzi M, et al. (2013) Receptor for Advanced Glycation Endproducts is upregulated in temporal lobe epilepsy and contributes to experimental seizures. Neurobiol Dis 58: 102-114. doi: 10.1016/j.nbd.2013.03.006
|
| [187] |
Maroso M, Balosso S, Ravizza T, et al. (2011) Interleukin-1 type 1 receptor/Toll-like receptor signalling in epilepsy: the importance of IL-1beta and high-mobility group box 1. J Intern Med 270: 319-326. doi: 10.1111/j.1365-2796.2011.02431.x
|
| [188] |
Vezzani A, Maroso M, Balosso S, et al. (2011) IL-1 receptor/Toll-like receptor signaling in infection, inflammation, stress and neurodegeneration couples hyperexcitability and seizures. Brain Behav Immun 25: 1281-1289. doi: 10.1016/j.bbi.2011.03.018
|
| [189] |
Zurolo E, Iyer A, Maroso M, et al. (2011) Activation of Toll-like receptor, RAGE and HMGB1 signalling in malformations of cortical development. Brain 134: 1015-1032. doi: 10.1093/brain/awr032
|
| [190] |
Gelderblom M, Sobey CG, Kleinschnitz C, et al. (2015) Danger signals in stroke. Ageing Res Rev 24: 77-82. doi: 10.1016/j.arr.2015.07.004
|
| [191] |
Gelderblom M, Leypoldt F, Steinbach K, et al. (2009) Temporal and spatial dynamics of cerebral immune cell accumulation in stroke. Stroke 40: 1849-1857. doi: 10.1161/STROKEAHA.108.534503
|
| [192] |
Kleinschnitz C, Kraft P, Dreykluft A, et al. (2013) Regulatory T cells are strong promoters of acute ischemic stroke in mice by inducing dysfunction of the cerebral microvasculature. Blood 121: 679-691. doi: 10.1182/blood-2012-04-426734
|
| [193] | Zhou Y, Xiong KL, Lin S, et al. (2010) Elevation of high-mobility group protein box-1 in serum correlates with severity of acute intracerebral hemorrhage. Mediators Inflamm 2010. |
| [194] |
Goldstein RS, Gallowitsch-Puerta M, Yang L, et al. (2006) Elevated high-mobility group box 1 levels in patients with cerebral and myocardial ischemia. Shock 25: 571-574. doi: 10.1097/01.shk.0000209540.99176.72
|
| [195] |
Vogelgesang A, May VE, Grunwald U, et al. (2010) Functional status of peripheral blood T-cells in ischemic stroke patients. PLoS One 5: e8718. doi: 10.1371/journal.pone.0008718
|
| [196] | Ramesh G, MacLean AG, Philipp MT (2013) Cytokines and chemokines at the crossroads of neuroinflammation, neurodegeneration, and neuropathic pain. Mediators Inflamm 2013. |
| [197] |
Moalem G, Tracey DJ (2006) Immune and inflammatory mechanisms in neuropathic pain. Brain Res Rev 51: 240-264. doi: 10.1016/j.brainresrev.2005.11.004
|
| [198] |
Coull JA, Beggs S, Boudreau D, et al. (2005) BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 438: 1017-1021. doi: 10.1038/nature04223
|
| [199] |
Miyoshi K, Obata K, Kondo T, et al. (2008) Interleukin-18-mediated microglia/astrocyte interaction in the spinal cord enhances neuropathic pain processing after nerve injury. J Neurosci 28: 12775-12787. doi: 10.1523/JNEUROSCI.3512-08.2008
|
| [200] |
Binshtok AM, Wang H, Zimmermann K, et al. (2008) Nociceptors are interleukin-1beta sensors. J Neurosci 28: 14062-14073. doi: 10.1523/JNEUROSCI.3795-08.2008
|
| [201] |
Skoff AM, Zhao C, Adler JE (2009) Interleukin-1alpha regulates substance P expression and release in adult sensory neurons. Exp Neurol 217: 395-400. doi: 10.1016/j.expneurol.2009.03.022
|
| [202] |
Xu XJ, Hao JX, Andell-Jonsson S, et al. (1997) Nociceptive responses in interleukin-6-deficient mice to peripheral inflammation and peripheral nerve section. Cytokine 9: 1028-1033. doi: 10.1006/cyto.1997.0243
|
| [203] |
Dominguez E, Mauborgne A, Mallet J, et al. (2010) SOCS3-mediated blockade of JAK/STAT3 signaling pathway reveals its major contribution to spinal cord neuroinflammation and mechanical allodynia after peripheral nerve injury. J Neurosci 30: 5754-5766. doi: 10.1523/JNEUROSCI.5007-09.2010
|
| [204] | Kim CF, Moalem-Taylor G (2011) Interleukin-17 contributes to neuroinflammation and neuropathic pain following peripheral nerve injury in mice. J Pain 12: 370-383. |
| [205] | Feldman P, Due MR, Ripsch MS, et al. (2012) The persistent release of HMGB1 contributes to tactile hyperalgesia in a rodent model of neuropathic pain. J Neuroinflammation 9: 180. |
| [206] |
Kuang X, Huang Y, Gu HF, et al. (2012) Effects of intrathecal epigallocatechin gallate, an inhibitor of Toll-like receptor 4, on chronic neuropathic pain in rats. Eur J Pharmacol 676: 51-56. doi: 10.1016/j.ejphar.2011.11.037
|
| [207] |
Maeda S, Hikiba Y, Shibata W, et al. (2007) Essential roles of high-mobility group box 1 in the development of murine colitis and colitis-associated cancer. Biochem Biophys Res Commun 360: 394-400. doi: 10.1016/j.bbrc.2007.06.065
|
| [208] |
Otoshi K, Kikuchi S, Kato K, et al. (2011) Anti-HMGB1 neutralization antibody improves pain-related behavior induced by application of autologous nucleus pulposus onto nerve roots in rats. Spine (Phila Pa 1976) 36: E692-698. doi: 10.1097/BRS.0b013e3181ecd675
|
| [209] |
Shibasaki M, Sasaki M, Miura M, et al. (2010) Induction of high mobility group box-1 in dorsal root ganglion contributes to pain hypersensitivity after peripheral nerve injury. Pain 149: 514-521. doi: 10.1016/j.pain.2010.03.023
|
| [210] | Li W, Li J, Sama AE, et al. (2013) Carbenoxolone blocks endotoxin-induced protein kinase R (PKR) activation and high mobility group box 1 (HMGB1) release. Mol Med 19: 203-211. |
| [211] |
Devi LA (2000) G-protein-coupled receptor dimers in the lime light. Trends Pharmacol Sci 21: 324-326. doi: 10.1016/S0165-6147(00)01519-4
|
| [212] |
Hanisch UK, Prinz M, Angstwurm K, et al. (2001) The protein tyrosine kinase inhibitor AG126 prevents the massive microglial cytokine induction by pneumococcal cell walls. Eur J Immunol 31: 2104-2115. doi: 10.1002/1521-4141(200107)31:7<2104::AID-IMMU2104>3.0.CO;2-3
|
| [213] |
Rock RB, Gekker G, Hu S, et al. (2004) Role of microglia in central nervous system infections. Clin Microbiol Rev 17: 942-964. doi: 10.1128/CMR.17.4.942-964.2004
|
| [214] |
Saukkonen K, Sande S, Cioffe C, et al. (1990) The role of cytokines in the generation of inflammation and tissue damage in experimental gram-positive meningitis. J Exp Med 171: 439-448. doi: 10.1084/jem.171.2.439
|
| [215] |
Zwijnenburg PJ, van der Poll T, Florquin S, et al. (2003) Interleukin-10 negatively regulates local cytokine and chemokine production but does not influence antibacterial host defense during murine pneumococcal meningitis. Infect Immun 71: 2276-2279. doi: 10.1128/IAI.71.4.2276-2279.2003
|
| [216] |
Ostergaard C, Yieng-Kow R, Larsen C, et al. (2000) Treatment with a monocolonal antibody to IL-8 attenuates the pleocytosis in experimental pneumococcal meningitis in rabbits when given intravenously, but not intracisternally. Clin Exp Immunol 122: 207-211. doi: 10.1046/j.1365-2249.2000.01357.x
|
| [217] |
Kielian T, Barry B, Hickey WF (2001) CXC Chemokine Receptor-2 Ligands Are Required for Neutrophil-Mediated Host Defense in Experimental Brain Abscesses1. J Immunol 166: 4634-4643. doi: 10.4049/jimmunol.166.7.4634
|
| [218] |
Kielian T, Phulwani NK, Esen N, et al. (2007) MyD88-dependent signals are essential for the host immune response in experimental brain abscess. J Immunol 178: 4528-4537. doi: 10.4049/jimmunol.178.7.4528
|
| [219] |
Kielian T, Mayes P, Kielian M (2002) Characterization of microglial responses to Staphylococcus aureus: effects on cytokine, costimulatory molecule, and Toll-like receptor expression. J Neuroimmunol 130: 86-99. doi: 10.1016/S0165-5728(02)00216-3
|
| [220] | Kielian T, Esen N, Bearden ED (2005) Toll‐like receptor 2 (TLR2) is pivotal for recognition of S. aureus peptidoglycan but not intact bacteria by microglia. Glia 49: 567-576. |
| [221] | Gurley C, Nichols J, Liu S, et al. (2008) Microglia and astrocyte activation by toll-like receptor ligands: modulation by PPAR-agonists. PPAR Res 2008. |
| [222] |
Esen N, Kielian T (2006) Central role for MyD88 in the responses of microglia to pathogen-associated molecular patterns. J Immunol 176: 6802-6811. doi: 10.4049/jimmunol.176.11.6802
|
| [223] |
Baldwin AC, Kielian T (2004) Persistent immune activation associated with a mouse model of Staphylococcus aureus-induced experimental brain abscess. J Neuroimmunol 151: 24-32. doi: 10.1016/j.jneuroim.2004.02.002
|
| [224] |
Tang D, Kang R, Cao L, et al. (2008) A pilot study to detect high mobility group box 1 and heat shock protein 72 in cerebrospinal fluid of pediatric patients with meningitis. Crit Care Med 36: 291-295. doi: 10.1097/01.CCM.0000295316.86942.CE
|
| [225] |
Asano T, Ichiki K, Koizumi S, et al. (2011) High mobility group box 1 in cerebrospinal fluid from several neurological diseases at early time points. Int J Neurosci 121: 480-484. doi: 10.3109/00207454.2011.580868
|
| [226] |
Hohne C, Wenzel M, Angele B, et al. (2013) High mobility group box 1 prolongs inflammation and worsens disease in pneumococcal meningitis. Brain 136: 1746-1759. doi: 10.1093/brain/awt064
|
| [227] |
Hemmi H, Takeuchi O, Kawai T, et al. (2000) A Toll-like receptor recognizes bacterial DNA. Nature 408: 740-745. doi: 10.1038/35047123
|
| [228] | Koedel U (2009) Toll-like receptors in bacterial meningitis. Toll-like Receptors: Roles in Infection and Neuropathology. Springer. pp. 15-40. |
| [229] |
Sanders MS, van Well GT, Ouburg S, et al. (2012) Toll-like receptor 9 polymorphisms are associated with severity variables in a cohort of meningococcal meningitis survivors. BMC Infect Dis 12: 112. doi: 10.1186/1471-2334-12-112
|
| [230] |
Alleva LM, Yang H, Tracey KJ, et al. (2005) High mobility group box 1 (HMGB1) protein: possible amplification signal in the pathogenesis of falciparum malaria. Trans R Soc Trop Med Hyg 99: 171-174. doi: 10.1016/j.trstmh.2004.06.008
|
| [231] |
Mostofa AG, Punganuru SR, Madala HR, et al. (2017) The Process and Regulatory Components of Inflammation in Brain Oncogenesis. Biomolecules 7: 34. doi: 10.3390/biom7020034
|
| [232] | Albulescu R, Codrici E, Popescu ID, et al. (2013) Cytokine patterns in brain tumour progression. Mediators Inflamm 2013: 979748. |
| [233] | Goswami S, Gupta A, Sharma SK (1998) Interleukin-6-mediated autocrine growth promotion in human glioblastoma multiforme cell line U87MG. J Neurochem 71: 1837-1845. |
| [234] |
Brat DJ, Bellail AC, Van Meir EG (2005) The role of interleukin-8 and its receptors in gliomagenesis and tumoral angiogenesis. Neuro Oncol 7: 122-133. doi: 10.1215/S1152851704001061
|
| [235] |
Yoshida S, Ono M, Shono T, et al. (1997) Involvement of interleukin-8, vascular endothelial growth factor, and basic fibroblast growth factor in tumor necrosis factor alpha-dependent angiogenesis. Mol Cell Biol 17: 4015-4023. doi: 10.1128/MCB.17.7.4015
|
| [236] | Tanabe K, Matsushima-Nishiwaki R, Yamaguchi S, et al. (2010) Mechanisms of tumor necrosis factor-alpha-induced interleukin-6 synthesis in glioma cells. J Neuroinflammation 7: 16. |
| [237] |
Ghosh S, Paul A, Sen E (2013) Tumor Necrosis Factor Alpha-Induced Hypoxia-Inducible Factor 1α–β-Catenin Axis Regulates Major Histocompatibility Complex Class I Gene Activation through Chromatin Remodeling. Mol Cell Biol 33: 2718-2731. doi: 10.1128/MCB.01254-12
|
| [238] |
Hao C, Parney IF, Roa WH, et al. (2002) Cytokine and cytokine receptor mRNA expression in human glioblastomas: evidence of Th1, Th2 and Th3 cytokine dysregulation. Acta Neuropathol 103: 171-178. doi: 10.1007/s004010100448
|
| [239] |
Grauer O, Poschl P, Lohmeier A, et al. (2007) Toll-like receptor triggered dendritic cell maturation and IL-12 secretion are necessary to overcome T-cell inhibition by glioma-associated TGF-beta2. J Neurooncol 82: 151-161. doi: 10.1007/s11060-006-9274-2
|
| [240] |
Calandra T, Roger T (2003) Macrophage migration inhibitory factor: a regulator of innate immunity. Nat Rev Immunol 3: 791-800. doi: 10.1038/nri1200
|
| [241] |
Mantovani A, Allavena P, Sica A, et al. (2008) Cancer-related inflammation. Nature 454: 436-444. doi: 10.1038/nature07205
|
| [242] |
Apte RN, Dotan S, Elkabets M, et al. (2006) The involvement of IL-1 in tumorigenesis, tumor invasiveness, metastasis and tumor-host interactions. Cancer Metastasis Rev 25: 387-408. doi: 10.1007/s10555-006-9004-4
|
| [243] |
Martins I, Kepp O, Menger L, et al. (2013) Fluorescent biosensors for the detection of HMGB1 release. Methods Mol Biol 1004: 43-56. doi: 10.1007/978-1-62703-383-1_4
|
| [244] | Zhang J, Liu C, Hou R (2014) Knockdown of HMGB1 improves apoptosis and suppresses proliferation and invasion of glioma cells. Chin J Cancer Res 26: 658-668. |
| [245] |
Kostova N, Zlateva S, Ugrinova I, et al. (2010) The expression of HMGB1 protein and its receptor RAGE in human malignant tumors. Mol Cell Biochem 337: 251-258. doi: 10.1007/s11010-009-0305-0
|
| [246] | Seidu RA, Wu M, Su Z, et al. (2017) Paradoxical Role of High Mobility Group Box 1 in Glioma: A Suppressor or a Promoter? Oncol Rev 11: 325. |
| [247] |
Jube S, Rivera ZS, Bianchi ME, et al. (2012) Cancer cell secretion of the DAMP protein HMGB1 supports progression in malignant mesothelioma. Cancer Res 72: 3290-3301. doi: 10.1158/0008-5472.CAN-11-3481
|
| [248] |
Yang GL, Zhang LH, Bo JJ, et al. (2012) Increased expression of HMGB1 is associated with poor prognosis in human bladder cancer. J Surg Oncol 106: 57-61. doi: 10.1002/jso.23040
|
| [249] |
Kim JB, Lim CM, Yu YM, et al. (2008) Induction and subcellular localization of high‐mobility group box–1 (HMGB1) in the postischemic rat brain. J Neurosci Res 86: 1125-1131. doi: 10.1002/jnr.21555
|
| [250] |
Jones KA, Thomsen C (2013) The role of the innate immune system in psychiatric disorders. Mol Cell Neurosci 53: 52-62. doi: 10.1016/j.mcn.2012.10.002
|
| [251] |
Frank MG, Weber MD, Watkins LR, et al. (2016) Stress-induced neuroinflammatory priming: A liability factor in the etiology of psychiatric disorders. Neurobiol Stress 4: 62-70. doi: 10.1016/j.ynstr.2015.12.004
|
| [252] |
Dantzer R, O'Connor JC, Freund GG, et al. (2008) From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci 9: 46-56. doi: 10.1038/nrn2297
|
| [253] |
Johnson JD, Campisi J, Sharkey CM, et al. (2005) Catecholamines mediate stress-induced increases in peripheral and central inflammatory cytokines. Neuroscience 135: 1295-1307. doi: 10.1016/j.neuroscience.2005.06.090
|
| [254] |
Weber MD, Frank MG, Sobesky JL, et al. (2013) Blocking toll-like receptor 2 and 4 signaling during a stressor prevents stress-induced priming of neuroinflammatory responses to a subsequent immune challenge. Brain Behav Immun 32: 112-121. doi: 10.1016/j.bbi.2013.03.004
|
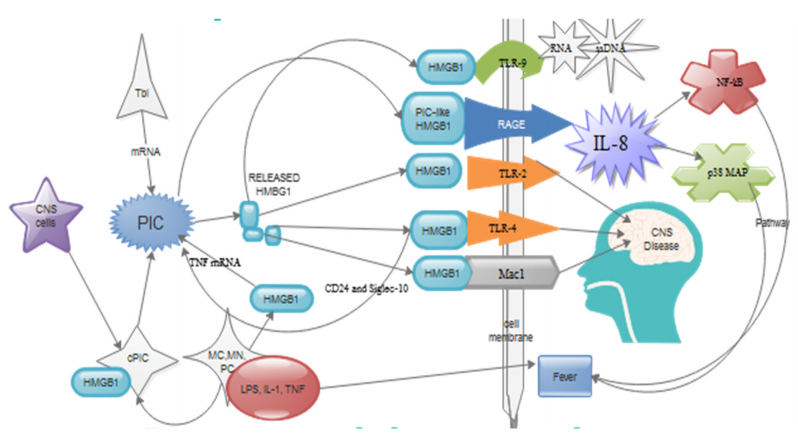
Figures(1) / Tables(1)

Seidu A. Richard, Wu Min, Zhaoliang Su, Hua-Xi Xu. Epochal neuroinflammatory role of high mobility group box 1 in central nervous system diseases[J]. AIMS Molecular Science, 2017, 4(2): 185-218. doi: 10.3934/molsci.2017.2.185







 DownLoad:
DownLoad: