Citation: Bashir Sajo Mienda, Faezah Mohd Salleh. Bio-succinic acid production: Escherichia coli strains design from genome-scale perspectives[J]. AIMS Bioengineering, 2017, 4(4): 418-430. doi: 10.3934/bioeng.2017.4.418

| [1] |
Zeikus JG, Jain MK, Elankovan P (1999) Biotechnology of succinic acid production and markets for derived industrial products. Appl Microbiol Biot 51: 545–552. doi: 10.1007/s002530051431

|
| [2] |
Forster AH, Gescher J (2014) Metabolic engineering of Escherichia coli for production of mixed-acid fermentation end products. Front Bioeng Biotechnol 2: 16. doi: 10.3389/fbioe.2014.00016
|
| [3] |
Cao Y, Zhang R, Sun C, et al. (2013) Fermentative succinate production: an emerging technology to replace the traditional petrochemical processes. Biomed Res Int 2013: 723412. doi: 10.1155/2013/723412
|
| [4] |
Sánchez AM, Bennett GN, San KY (2005) Novel pathway engineering design of the anaerobic central metabolic pathway in Escherichia coli to increase succinate yield and productivity. Metab Eng 7: 229–239. doi: 10.1016/j.ymben.2005.03.001
|
| [5] |
Thakker C, Martinez I, San KY, et al. (2012) Succinate production in Escherichia coli. Biotechnol J 7: 213–224. doi: 10.1002/biot.201100061
|
| [6] |
Sanchez AM, Bennett GN, San KY (2005) Efficient succinic acid production from glucose through overexpression of pyruvate carboxylase in an Escherichia coli alcohol dehydrogenase and lactate dehydrogenase mutant. Biotechnol Progr 21: 358–365. doi: 10.1021/bp049676e
|
| [7] |
Cox SJ, Shalel LS, Sanchez A, et al. (2006) Development of a metabolic network design and optimization framework incorporating implementation constraints: a succinate production case study. Metab Eng 8: 46–57. doi: 10.1016/j.ymben.2005.09.006
|
| [8] |
Lin H, Bennett GN, San KY (2005) Metabolic engineering of aerobic succinate production systems in Escherichia coli to improve process productivity and achieve the maximum theoretical succinate yield. Metab Eng 7: 116–127. doi: 10.1016/j.ymben.2004.10.003
|
| [9] |
Feist AM, Henry CS, Reed JL, et al. (2007) A genome-scale metabolic reconstruction for Escherichia coli K-12 MG1655 that accounts for 1260 ORFs and thermodynamic information. Mol Syst Biol 3: 121–138. doi: 10.1038/msb4100155
|
| [10] |
Orth JD, Conrad TM, Na J, et al. (2011) A comprehensive genome-scale reconstruction of Escherichia coli metabolism-2011. Mol Syst Biol 7: 535–543. doi: 10.1038/msb.2011.65
|
| [11] |
Feist AM, Zielinski DC, Orth JD, et al. (2010) Model-driven evaluation of the production potential for growth-coupled products of Escherichia coli. Metab Eng 12: 173–186. doi: 10.1016/j.ymben.2009.10.003
|
| [12] |
King ZA, O'Brien EJ, Feist AM, et al. (2017) Literature mining supports a next-generation modeling approach to predict cellular byproduct secretion. Metab Eng 39: 220–227. doi: 10.1016/j.ymben.2016.12.004
|
| [13] |
McCloskey D, Palsson BO, Feist AM (2013) Basic and applied uses of genome-scale metabolic network reconstructions of Escherichia coli. Mol Syst Biol 9: 661–675. doi: 10.1038/msb.2013.18
|
| [14] |
Mienda BS, Shamsir MS, Illias RM (2016) Model-guided metabolic gene knockout of gnd for enhanced succinate production in Escherichia coli from glucose and glycerol substrates. Comput Biol Chem 61: 130–137. doi: 10.1016/j.compbiolchem.2016.01.013
|
| [15] |
Mienda BS, Shamsir MS, Md IR (2015) Model-aided atpE gene knockout strategy in Escherichia coli for enhanced succinic acid production from glycerol. J Biomol Struct Dyn 34: 1705–1716. doi: 10.1080/07391102.2015.1090341
|
| [16] |
Mienda BS, Shamsir MS, Md IR (2015) Model-assisted formate dehydrogenase-O (fdoH) gene knockout for enhanced succinate production in Escherichia coli from glucose and glycerol carbon sources. J Biomol Struct Dyn 34: 2305–2316. doi: 10.1080/07391102.2015.1113387
|
| [17] |
Lee SY, Kim HU (2015) Systems strategies for developing industrial microbial strains. Nat Biotechnol 33: 1061–1072. doi: 10.1038/nbt.3365
|
| [18] |
Lee SJ, Song H, Lee SY (2006) Genome-based metabolic engineering of Mannheimia succiniciproducens for succinic acid production. Appl Environ Microb 72: 1939–1948. doi: 10.1128/AEM.72.3.1939-1948.2006
|
| [19] |
Becker J, Reinefeld J, Stellmacher R, et al. (2013) Systems-wide analysis and engineering of metabolic pathway fluxes in bio-succinate producing Basfia succiniciproducens. Biotechnol Bioeng 110: 3013–3023. doi: 10.1002/bit.24963
|
| [20] |
Park SH, Kim HU, Kim TY, et al. (2014) Metabolic engineering of Corynebacterium glutamicum for L-arginine production. Nat Commun 5: 4618–4626. doi: 10.1038/ncomms5618
|
| [21] |
Becker J, Zelder O, Hafner S, et al. (2011) From zero to hero-design-based systems metabolic engineering of Corynebacterium glutamicum for L-lysine production. Metab Eng 13: 159–168. doi: 10.1016/j.ymben.2011.01.003
|
| [22] |
Kind S, Neubauer S, Becker J, et al. (2014) From zero to hero-production of bio-based nylon from renewable resources using engineered Corynebacterium glutamicum. Metab Eng 25: 113–123. doi: 10.1016/j.ymben.2014.05.007
|
| [23] |
Choi S, Song CW, Shin JH, et al. (2015) Biorefineries for the production of top building block chemicals and their derivatives. Metab Eng 28: 223–239. doi: 10.1016/j.ymben.2014.12.007
|
| [24] |
Chung H, Yang JE, Ha JY, et al. (2015) Bio-based production of monomers and polymers by metabolically engineered microorganisms. Curr Opin Biotech 36: 73–84. doi: 10.1016/j.copbio.2015.07.003
|
| [25] |
Yin X, Li J, Shin Hd, et al. (2015) Metabolic engineering in the biotechnological production of organic acids in the tricarboxylic acid cycle of microorganisms: advances and prospects. Biotechnol Adv 33: 830. doi: 10.1016/j.biotechadv.2015.04.006
|
| [26] |
Monk J, Palsson BO (2014) Genetics. Predicting microbial growth. Science 344: 1448–1449. doi: 10.1126/science.1253388
|
| [27] |
Kim B, Kim WJ, Kim DI, et al. (2015) Applications of genome-scale metabolic network model in metabolic engineering. J Ind Microbiol Biot 42: 339–348. doi: 10.1007/s10295-014-1554-9
|
| [28] |
Le NN, Bornstein B, Broicher A, et al. (2006) BioModels database: a free, centralized database of curated, published, quantitative kinetic models of biochemical and cellular systems. Nucleic Acids Res 34: D689–D691. doi: 10.1093/nar/gkj092
|
| [29] |
Schellenberger J, Park JO, Conrad TM, et al. (2010) BiGG: a biochemical genetic and genomic knowledgebase of large scale metabolic reconstructions. BMC Bioinformatics 11: 213–222. doi: 10.1186/1471-2105-11-213
|
| [30] |
O'Brien EJ, Lerman JA, Chang RL, et al. (2013) Genome-scale models of metabolism and gene expression extend and refine growth phenotype prediction. Mol Syst Biol 9: 693–693. doi: 10.1038/msb.2013.52
|
| [31] |
Guzman GI, Utrilla J, Nurk S, et al. (2015) Model-driven discovery of underground metabolic functions in Escherichia coli. Proc Natl Acad Sci USA 112: 929–934. doi: 10.1073/pnas.1414218112
|
| [32] |
Mienda BS (2017) Genome-scale metabolic models as platforms for strain design and biological discovery. J Biomol Struct Dyn 35: 1863–1873. doi: 10.1080/07391102.2016.1197153
|
| [33] | Mienda BS, Shamsir MS (2015) Model-guided in silico deletion of pntA gene predicts increased succinate production under anaerobic conditions in Escherichia coli. Biosci Bioeng Commun 1: 1–10. |
| [34] |
Mienda BS, Shahir SM (2015) Model-driven in silico glpc gene knockout predicts increased succinate production from glycerol in Escherichia Coli. AIMS Bioeng 2: 40–48. doi: 10.3934/bioeng.2015.2.40
|
| [35] |
Mienda BS, Shamsir MS (2015) In silico deletion of PtsG gene in Escherichia coli genome-scale model predicts increased succinate production from glycerol. J Biomol Struct Dyn 33: 2380–2389. doi: 10.1080/07391102.2015.1036461
|
| [36] |
Mienda BS, Shamsir MS (2015) Model-aided anaerobic metabolic gene knockout of malate dehydrogenase (mdh) gene predicts increased succinate production in Escherichia coli. J Biomol Struct Dyn 33: 98–100. doi: 10.1080/07391102.2015.1032786
|
| [37] |
Lee SJ, Lee DY, Kim TY, et al. (2005) Metabolic engineering of Escherichia coli for enhanced production of succinic acid, based on genome comparison and in silico gene knockout simulation. Appl Environ Microbiol 71: 7880–7887. doi: 10.1128/AEM.71.12.7880-7887.2005
|
| [38] |
Jantama K, Zhang X, Moore JC, et al. (2008) Eliminating side products and increasing succinate yields in engineered strains of Escherichia coli C. Biotechnol Bioeng 101: 881–893. doi: 10.1002/bit.22005
|
| [39] |
Kim TY, Park JM, Kim HU, et al. (2015) Design of homo-organic acid producing strains using multi-objective optimization. Metab Eng 28: 63–73. doi: 10.1016/j.ymben.2014.11.012
|
| [40] |
Yim H, Haselbeck R, Niu W, et al. (2011) Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nat Chem Biol 7: 445–452. doi: 10.1038/nchembio.580
|
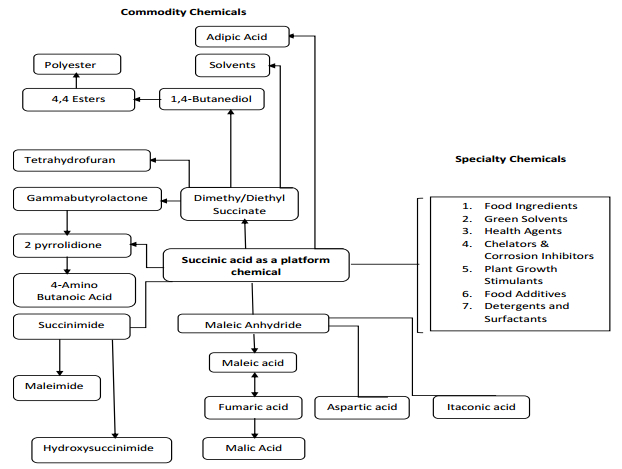
Figures(1) / Tables(3)

Bashir Sajo Mienda, Faezah Mohd Salleh. Bio-succinic acid production: Escherichia coli strains design from genome-scale perspectives[J]. AIMS Bioengineering, 2017, 4(4): 418-430. doi: 10.3934/bioeng.2017.4.418







 DownLoad:
DownLoad: