Citation: Ling Wang, Shunbin Ning. “Toll-free” pathways for production of type I interferons[J]. AIMS Allergy and Immunology, 2017, 1(3): 143-163. doi: 10.3934/Allergy.2017.3.143

| [1] |
Takaoka A, Wang Z, Choi MK, et al. (2007) DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 448: 501–505. doi: 10.1038/nature06013

|
| [2] |
Kim T, Pazhoor S, Bao M, et al. (2010) Aspartate-glutamate-alanine-histidine box motif (DEAH)/RNA helicase A helicases sense microbial DNA in human plasmacytoid dendritic cells. Proc Natl Acad Sci USA 107: 15181–15186. doi: 10.1073/pnas.1006539107
|
| [3] |
Parvatiyar K, Zhang Z, Teles RM, et al. (2012) The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nat Immunol 13: 1155–1161. doi: 10.1038/ni.2460
|
| [4] |
Zhang Z, Yuan B, Bao M, et al. (2011) The helicase DDX41 senses intracellular DNA mediated by the adaptor STING in dendritic cells. Nat Immunol 12: 959–965. doi: 10.1038/ni.2091
|
| [5] |
Chiu YH, MacMillan JB, Chen ZJ (2009) RNA polymerase III detects cytosolic DNA and induces type I interferons through the RIG-I pathway. Cell 138: 576–591. doi: 10.1016/j.cell.2009.06.015
|
| [6] |
Zhang X, Brann TW, Zhou M, et al. (2011) Ku70 is a novel cytosolic DNA sensor that induces type-III rather than type-I IFN. J Immunol 186: 4541–4545. doi: 10.4049/jimmunol.1003389
|
| [7] |
Kondo T, Kobayashi J, Saitoh T, et al. (2013) DNA damage sensor MRE11 recognizes cytosolic double-stranded DNA and induces type I interferon by regulating STING trafficking. Proc Natl Acad Sci USA 110: 2969–2974. doi: 10.1073/pnas.1222694110
|
| [8] |
Xia P, Wang S, Ye B, et al. (2015) Sox2 functions as a sequence-specific DNA sensor in neutrophils to initiate innate immunity against microbial infection. Nat Immunol 16: 366–375. doi: 10.1038/ni.3117
|
| [9] |
Yang P, An H, Liu X, et al. (2010) The cytosolic nucleic acid sensor LRRFIP1 mediates the production of type I interferon via a beta-catenin-dependent pathway. Nat Immunol 11: 487–494. doi: 10.1038/ni.1876
|
| [10] | Pichlmair A, Lassnig C, Eberle CA, et al. (2011) IFIT1 is an antiviral protein that recognizes 5'-triphosphate RNA. Nat Immunol 12: 624–630. |
| [11] |
Hornung V, Hartmann R, Ablasser A, et al. (2014) OAS proteins and cGAS: unifying concepts in sensing and responding to cytosolic nucleic acids. Nat Rev Immunol 14: 521–528. doi: 10.1038/nri3719
|
| [12] |
Ori D, Murase M, Kawai T (2017) Cytosolic nucleic acid sensors and innate immune regulation. Int Rev Immunol 36: 74–88. doi: 10.1080/08830185.2017.1298749
|
| [13] |
Xia P, Wang S, Gao P, et al. (2016) DNA sensor cGAS-mediated immune recognition. Protein Cell 7: 777–791. doi: 10.1007/s13238-016-0320-3
|
| [14] |
Schlee M, Hartmann G (2016) Discriminating self from non-self in nucleic acid sensing. Nat Rev Immunol 16: 566–580. doi: 10.1038/nri.2016.78
|
| [15] |
Ning S, Pagano J, Barber G (2011) IRF7: activation, regulation, modification, and function. Genes Immun 12: 399–414. doi: 10.1038/gene.2011.21
|
| [16] |
Chen Q, Sun L, Chen ZJ (2016) Regulation and function of the cGAS-STING pathway of cytosolic DNA sensing. Nat Immunol 17: 1142–1149. doi: 10.1038/ni.3558
|
| [17] |
Wilson EB, Yamada DH, Elsaesser H, et al. (2013) Blockade of chronic type I interferon signaling to control persistent LCMV infection. Science 340: 202–207. doi: 10.1126/science.1235208
|
| [18] |
Teijaro JR, Ng C, Lee AM, et al. (2013) Persistent LCMV infection is controlled by blockade of type I interferon signaling. Science 340: 207–211. doi: 10.1126/science.1235214
|
| [19] |
Cha L, Berry CM, Nolan D, et al. (2014) Interferon-alpha, immune activation and immune dysfunction in treated HIV infection. Clin Trans Immunol 3: e10. doi: 10.1038/cti.2014.1
|
| [20] |
Catalfamo M, Wilhelm C, Tcheung L, et al. (2011) CD4 and CD8 T cell immune activation during chronic HIV infection: roles of homeostasis, HIV, type I IFN, and IL-7. J Immunol 186: 2106–2116. doi: 10.4049/jimmunol.1002000
|
| [21] |
Crouse J, Kalinke U, Oxenius A (2015) Regulation of antiviral T cell responses by type I interferons. Nat Rev Immunol 15: 231–242. doi: 10.1038/nri3806
|
| [22] |
Bosque A, Planelles V (2009) Induction of HIV-1 latency and reactivation in primary memory CD4+ T cells. Blood 113: 58–65. doi: 10.1182/blood-2008-07-168393
|
| [23] |
Härtlova A, Erttmann SF, Raffi FAM, et al. (2015) DNA damage primes the type i interferon system via the cytosolic DNA sensor STING to promote anti-microbial innate immunity. Immunity 42: 332–343. doi: 10.1016/j.immuni.2015.01.012
|
| [24] |
White MJ, McArthur K, Metcalf D, et al. (2014) Apoptotic caspases suppress mtDNA-induced STING-mediated type I IFN production. Cell 159: 1549–1562. doi: 10.1016/j.cell.2014.11.036
|
| [25] |
Vilenchik MM, Knudson AG (2003) Endogenous DNA double-strand breaks: production, fidelity of repair, and induction of cancer. Proc Natl Acad Sci USA 100: 12871–12876. doi: 10.1073/pnas.2135498100
|
| [26] |
de Galarreta MR, Lujambio A (2017) DNA sensing in senescence. Nat Cell Biol 19: 1008–1009. doi: 10.1038/ncb3603
|
| [27] | Gluck S, Guey B, Gulen MF, et al. (2017) Innate immune sensing of cytosolic chromatin fragments through cGAS promotes senescence. Nat Cell Biol 19: In press. |
| [28] | Ng KW, Marshall EA, Bell JC, et al. (2017) cGAS-STING and cancer: dichotomous roles in tumor immunity and development. Trends Immunol: In press. |
| [29] |
Yang H, Wang H, Ren J, et al. (2017) cGAS is essential for cellular senescence. Proc Natl Acad Sci USA 114: E4612–E4620. doi: 10.1073/pnas.1705499114
|
| [30] |
Baccala R, Hoebe K, Kono DH, et al. (2007) TLR-dependent and TLR-independent pathways of type I interferon induction in systemic autoimmunity. Nat Med 13: 543–551. doi: 10.1038/nm1590
|
| [31] |
Shibutani ST, Saitoh T, Nowag H, et al. (2015) Autophagy and autophagy-related proteins in the immune system. Nat Immunol 16: 1014–1024. doi: 10.1038/ni.3273
|
| [32] |
Agod Z, Fekete T, Budai MM, et al. (2017) Regulation of type I interferon responses by mitochondria-derived reactive oxygen species in plasmacytoid dendritic cells. Redox Biol 13: 633–645. doi: 10.1016/j.redox.2017.07.016
|
| [33] |
McNab F, Mayer-Barber K, Sher A, et al. (2015) Type I interferons in infectious disease. Nat Rev Immunol 15: 87–103. doi: 10.1038/nri3787
|
| [34] |
Forster S (2012) Interferon signatures in immune disorders and disease. Immunol Cell Biol 90: 520–527. doi: 10.1038/icb.2012.12
|
| [35] |
Elkon KB, Wiedeman A (2012) Type I IFN system in the development and manifestations of SLE. Curr Opin Rheumatol 24: 499–505. doi: 10.1097/BOR.0b013e3283562c3e
|
| [36] |
Sandler NG, Bosinger SE, Estes JD, et al. (2014) Type I interferon responses in rhesus macaques prevent SIV infection and slow disease progression. Nature 511: 601–605. doi: 10.1038/nature13554
|
| [37] |
Mogensen T, Melchjorsen J, Larsen C, et al. (2010) Innate immune recognition and activation during HIV infection. Retrovirology 7: 54. doi: 10.1186/1742-4690-7-54
|
| [38] |
Tough DF (2012) Modulation of T-cell function by type I interferon. Immunol Cell Biol 90: 492–497. doi: 10.1038/icb.2012.7
|
| [39] |
Zitvogel L, Galluzzi L, Kepp O, et al. (2015) Type I interferons in anticancer immunity. Nat Rev Immunol 15: 405–414. doi: 10.1038/nri3845
|
| [40] |
Gajewski TF, Corrales L (2015) New perspectives on type I IFNs in cancer. Cytokine Growth F R 26: 175–178. doi: 10.1016/j.cytogfr.2015.01.001
|
| [41] | Dominguez-Villar M, Gautron AS, de Marcken M, et al. (2015) TLR7 induces anergy in human CD4+ T cells. Nat Immunol 16: 118–128. |
| [42] | Andreeva L, Hiller B, Kostrewa D, et al. (2017) cGAS senses long and HMGB/TFAM-bound U-turn DNA by forming protein-DNA ladders. Nature: In press. |
| [43] |
Cordaux R, Batzer MA (2009) The impact of retrotransposons on human genome evolution. Nat Rev Genet 10: 691–703. doi: 10.1038/nrg2640
|
| [44] | Liu S, Cai X, Wu J, et al. (2015) Phosphorylation of innate immune adaptor proteins MAVS, STING, and TRIF induces IRF3 activation. Science 347. |
| [45] |
Wang Q, Liu X, Cui Y, et al. (2014) The E3 ubiquitin ligase AMFR and INSIG1 bridge the activation of TBK1 kinase by modifying the adaptor STING. Immunity 41: 919–933. doi: 10.1016/j.immuni.2014.11.011
|
| [46] |
Man SM, Karki R, Kanneganti TD (2016) AIM2 inflammasome in infection, cancer, and autoimmunity: role in DNA sensing, inflammation, and innate immunity. Eur J Immunol 46: 269–280. doi: 10.1002/eji.201545839
|
| [47] |
Diner BA, Lum KK, Cristea IM (2015) The emerging role of nuclear viral DNA sensors. J Biol Chem 290: 26412–26421. doi: 10.1074/jbc.R115.652289
|
| [48] |
West AP, Khoury-Hanold W, Staron M, et al. (2015) Mitochondrial DNA stress primes the antiviral innate immune response. Nature 520: 553–557. doi: 10.1038/nature14156
|
| [49] |
Ansari MA, Singh VV, Dutta S, et al. (2013) Constitutive interferon-inducible protein 16-inflammasome activation during Epstein-Barr virus latency I, II, and III in B and epithelial cells. J Virol 87: 8606–8623. doi: 10.1128/JVI.00805-13
|
| [50] |
Christensen MH, Paludan SR (2017) Viral evasion of DNA-stimulated innate immune responses. Cell Mol Immunol 14: 4–13. doi: 10.1038/cmi.2016.06
|
| [51] |
Zevini A, Olagnier D, Hiscott J (2017) Crosstalk between cytoplasmic RIG-I and STING sensing pathways. Trends Immunol 38: 194–205. doi: 10.1016/j.it.2016.12.004
|
| [52] |
Satoh T, Kato H, Kumagai Y, et al. (2010) LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses. Proc Natl Acad Sci USA 107: 1512–1517. doi: 10.1073/pnas.0912986107
|
| [53] |
Kato K, Omura H, Ishitani R, et al. (2017) Cyclic GMP-AMP as an endogenous second messenger in innate immune signaling by cytosolic DNA. Annu Rev Biochem 86: 541–566. doi: 10.1146/annurev-biochem-061516-044813
|
| [54] |
Wu JJ, Li W, Shao Y, et al. (2015) Inhibition of cGAS DNA sensing by a herpesvirus virion protein. Cell Host Microbe 18: 333–344. doi: 10.1016/j.chom.2015.07.015
|
| [55] |
Li W, Avey D, Fu B, et al. (2016) Kaposi's sarcoma-associated herpesvirus inhibitor of cGAS (KicGAS), encoded by orf52, is an abundant tegument protein and is required for production of infectious progeny viruses. J Virol 90: 5329–5342. doi: 10.1128/JVI.02675-15
|
| [56] |
Ma Z, Jacobs SR, West JA, et al. (2015) Modulation of the cGAS-STING DNA sensing pathway by gammaherpesviruses. Proc Natl Acad Sci USA 112: E4306–E4315. doi: 10.1073/pnas.1503831112
|
| [57] |
Hwang SW, Kim D, Jung JU, et al. (2017) KSHV-encoded viral interferon regulatory factor 4 (vIRF4) interacts with IRF7 and inhibits interferon alpha production. Biochem Bioph Res Co 486: 700–705. doi: 10.1016/j.bbrc.2017.03.101
|
| [58] |
Lau L, Gray EE, Brunette RL, et al. (2015) DNA tumor virus oncogenes antagonize the cGAS-STING DNA-sensing pathway. Science 350: 568–571. doi: 10.1126/science.aab3291
|
| [59] | de Souza RF, Iyer LM, Aravind L (2010) Diversity and evolution of chromatin proteins encoded by DNA viruses. BBA-Gene Regul Mech 1799: 302–318. |
| [60] |
Towers GJ, Noursadeghi M (2014) Interactions between HIV-1 and the cell-autonomous innate immune system. Cell Host Microbe 16: 10–18. doi: 10.1016/j.chom.2014.06.009
|
| [61] | Sandstrom TS, Ranganath N, Angel JB (2017) Impairment of the type I interferon response by HIV-1: potential targets for HIV eradication. Cytokine Growth F R: In press. |
| [62] |
Rongvaux A, Jackson R, Harman CCD, et al. (2014) Apoptotic caspases prevent the induction of type I interferons by mitochondrial DNA. Cell 159: 1563–1577. doi: 10.1016/j.cell.2014.11.037
|
| [63] |
Zheng Q, Hou J, Zhou Y, et al. (2017) The RNA helicase DDX46 inhibits innate immunity by entrapping m6A-demethylated antiviral transcripts in the nucleus. Nat Immunol 18: 1094–1103. doi: 10.1038/ni.3830
|
| [64] | Boss IW, Renne R (2011) Viral miRNAs and immune evasion. BBA-Gene Regul Mech 1809: 708–714. |
| [65] |
Cullen BR (2013) MicroRNAs as mediators of viral evasion of the immune system. Nat Immunol 14: 205–210. doi: 10.1038/ni.2537
|
| [66] |
Wang L, Li G, Yao ZQ, et al. (2015) MicroRNA regulation of viral immunity, latency, and carcinogenesis of selected tumor viruses and HIV. Rev Med Virol 25: 320–341. doi: 10.1002/rmv.1850
|
| [67] |
Ding L, Huang XF, Dong GJ, et al. (2015) Activated STING enhances Tregs infiltration in the HPV-related carcinogenesis of tongue squamous cells via the c-jun/CCL22 signal. BBA-Mol Basis Dis 1852: 2494–2503. doi: 10.1016/j.bbadis.2015.08.011
|
| [68] |
Yarbrough ML, Zhang K, Sakthivel R, et al. (2014) Primate-specific miR-576-3p sets host defense signaling threshold. Nat Commun 5: 4963–4963. doi: 10.1038/ncomms5963
|
| [69] |
Wu MZ, Cheng WC, Chen SF, et al. (2017) miR-25/93 mediates hypoxia-induced immunosuppression by repressing cGAS. Nat Cell Biol 19: 1286–1296. doi: 10.1038/ncb3615
|
| [70] |
Yuan F, Dutta T, Wang L, et al. (2015) Human DNA exonuclease TREX1 is also an exoribonuclease that acts on single-stranded RNA. J Biol Chem 290: 13344–13353. doi: 10.1074/jbc.M115.653915
|
| [71] |
Samuel CE (2011) Adenosine deaminases acting on RNA (ADARs) are both antiviral and proviral dependent upon the virus. Virology 411: 180–193. doi: 10.1016/j.virol.2010.12.004
|
| [72] |
Liddicoat BJ, Piskol R, Chalk AM, et al. (2015) RNA editing by ADAR1 prevents MDA5 sensing of endogenous dsRNA as nonself. Science 349: 1115–1120. doi: 10.1126/science.aac7049
|
| [73] |
Yang S, Deng P, Zhu Z, et al. (2014) ADAR1 Limits RIG-I RNA detection and suppresses IFN production responding to viral and endogenous RNAs. J Immunol 193: 3436–3445. doi: 10.4049/jimmunol.1401136
|
| [74] |
Wang Q, Li X, Qi R, et al. (2017) RNA Editing, ADAR1, and the innate immune response. Genes 8: 41. doi: 10.3390/genes8010041
|
| [75] |
Gandy SZ, Linnstaedt SD, Muralidhar S, et al. (2007) RNA editing of the human herpesvirus 8 kaposin transcript eliminates its transforming activity and is induced during lytic replication. J Virol 81: 13544–13551. doi: 10.1128/JVI.01521-07
|
| [76] |
Iizasa H, Wulff BE, Alla NR, et al. (2010) Editing of Epstein-Barr virus-encoded BART6 microRNAs controls their dicer targeting and consequently affects viral latency. J Biol Chem 285: 33358–33370. doi: 10.1074/jbc.M110.138362
|
| [77] |
Rebhandl S, Huemer M, Greil R, et al. (2015) AID/APOBEC deaminases and cancer. Oncoscience 2: 320–333. doi: 10.18632/oncoscience.155
|
| [78] |
Konno H, Konno K, Barber GN (2013) Cyclic dinucleotides trigger ULK1 (ATG1) phosphorylation of STING to prevent sustained innate immune signaling. Cell 155: 688–698. doi: 10.1016/j.cell.2013.09.049
|
| [79] |
Seo GJ, Yang A, Tan B, et al. (2015) Akt kinase-mediated checkpoint of cGAS DNA sensing pathway. Cell Rep 13: 440–449. doi: 10.1016/j.celrep.2015.09.007
|
| [80] | Li S, Zhu M, Pan R, et al. (2016) The tumor suppressor PTEN has a critical role in antiviral innate immunity. Nat Immunol 17: 241–249. |
| [81] | Nekhai S, Jerebtsova M, Jackson A, et al. (2007) Regulation of HIV-1 transcription by protein phosphatase 1. Curr Hiv Res 5: 3–9. |
| [82] |
Wies E, Wang MK, Maharaj NP, et al. (2013) Dephosphorylation of the RNA sensors RIG-I and MDA5 by the phosphatase PP1 is essential for innate immune signaling. Immunity 38: 437–449. doi: 10.1016/j.immuni.2012.11.018
|
| [83] |
Davis ME, Wang MK, Rennick LJ, et al. (2014) Antagonism of the phosphatase PP1 by the measles virus V protein is required for innate immune escape of MDA5. Cell Host Microbe 16: 19–30. doi: 10.1016/j.chom.2014.06.007
|
| [84] |
Opaluch AM, Schneider M, Chiang CY, et al. (2014) Positive regulation of TRAF6-dependent innate immune responses by protein phosphatase PP1-gamma. Plos One 9: e89284. doi: 10.1371/journal.pone.0089284
|
| [85] |
Ilinykh PA, Tigabu B, Ivanov A, et al. (2014) Role of protein phosphatase 1 in dephosphorylation of Ebola virus VP30 protein and its targeting for the inhibition of viral transcription. J Biol Chem 289: 22723–22738. doi: 10.1074/jbc.M114.575050
|
| [86] | Cougot D, Allemand E, Rivière L, et al. (2012) Inhibition of PP1 phosphatase activity by HBx: a mechanism for the activation of hepatitis B virus transcription. Sci Signal 5: ra1. |
| [87] | Gu M, Zhang T, Lin W, et al. (2014) Protein phosphatase PP1 negatively regulates the Toll-like receptor- and RIG-I-like receptor-triggered production of type I interferon by inhibiting IRF3 phosphorylation at serines 396 and 385 in macrophage. Sci Signal: In press. |
| [88] |
Gu M, Ouyang C, Lin W, et al. (2014) Phosphatase holoenzyme PP1/GADD34 negatively regulates TLR response by inhibiting TAK1 serine 412 phosphorylation. J Immunol 192: 2846–2856. doi: 10.4049/jimmunol.1302537
|
| [89] | Clavarino G, Claudio N, Dalet A, et al. (2012) Protein phosphatase 1 subunit Ppp1r15a/GADD34 regulates cytokine production in polyinosinic: polycytidylic acid-stimulated dendritic cells. Proc Natl Acad Sci USA: In press. |
| [90] |
Peng D, Wang Z, Huang A, et al. (2017) A novel function of F-Box protein FBXO17 in negative regulation of type I IFN signaling by recruiting PP2A for IFN regulatory factor 3 deactivation. J Immunol 198: 808–819. doi: 10.4049/jimmunol.1601009
|
| [91] |
Shanker V, Trincucci G, Heim HM, et al. (2013) Protein phosphatase 2A impairs IFNα-induced antiviral activity against the hepatitis C virus through the inhibition of STAT1 tyrosine phosphorylation. J Viral Hepatitis 20: 612–621. doi: 10.1111/jvh.12083
|
| [92] |
Wang L, Zhao J, Ren J, et al. (2016) Protein phosphatase 1 abrogates IRF7-mediated type I IFN response in antiviral immunity. Eur J Immunol 46: 2409–2419. doi: 10.1002/eji.201646491
|
| [93] | Davis ME, Gack MU (2015) Ubiquitination in the antiviral immune response. Virology 479–480: 52–65. |
| [94] |
Lin D, Zhong B (2015) Regulation of cellular innate antiviral signaling by ubiquitin modification. Acta Biochim Biophys Sin (Shanghai) 47: 149–155. doi: 10.1093/abbs/gmu133
|
| [95] |
Heaton SM, Borg NA, Dixit VM (2016) Ubiquitin in the activation and attenuation of innate antiviral immunity. J Exp Med 213: 1–13. doi: 10.1084/jem.20151531
|
| [96] |
Zhou Y, He C, Lin W, et al. (2017) Post-translational regulation of antiviral innate signaling. Eur J Immunol 47: 1414–1426. doi: 10.1002/eji.201746959
|
| [97] |
van Tol S, Hage A, Giraldo M, et al. (2017) The TRIMendous role of TRIMs in virus-host interactions. Vaccines 5: 23. doi: 10.3390/vaccines5030023
|
| [98] |
Damgaard RB, Nachbur U, Yabal M, et al. (2012) The ubiquitin ligase XIAP recruits LUBAC for NOD2 signaling in inflammation and innate immunity. Mol Cell 46: 746–758. doi: 10.1016/j.molcel.2012.04.014
|
| [99] |
Keusekotten K, Elliott PR, Glockner L, et al. (2013) OTULIN antagonizes LUBAC signaling by specifically hydrolyzing Met1-linked polyubiquitin. Cell 153: 1312–1326. doi: 10.1016/j.cell.2013.05.014
|
| [100] |
Rivkin E, Almeida SM, Ceccarelli DF, et al. (2013) The linear ubiquitin-specific deubiquitinase gumby regulates angiogenesis. Nature 498: 318–324. doi: 10.1038/nature12296
|
| [101] |
Takiuchi T, Nakagawa T, Tamiya H, et al. (2014) Suppression of LUBAC-mediated linear ubiquitination by a specific interaction between LUBAC and the deubiquitinases CYLD and OTULIN. Genes Cells 19: 254–272. doi: 10.1111/gtc.12128
|
| [102] |
Tokunaga F, Nishimasu H, Ishitani R, et al. (2012) Specific recognition of linear polyubiquitin by A20 zinc finger 7 is involved in NFκB regulation. Embo J 31: 3856–3870. doi: 10.1038/emboj.2012.241
|
| [103] |
Hrdinka M, Fiil BK, Zucca M, et al. (2016) CYLD Limits Lys63- and Met1-Linked Ubiquitin at receptor complexes to regulate innate immune signaling. Cell Rep 14: 2846–2858. doi: 10.1016/j.celrep.2016.02.062
|
| [104] |
Damgaard RB, Walker JA, Marco-Casanova P, et al. (2016) The deubiquitinase OTULIN is an essential negative regulator of inflammation and autoimmunity. Cell 166: 1215–1230. doi: 10.1016/j.cell.2016.07.019
|
| [105] |
Wang Q, Huang L, Hong Z, et al. (2017) The E3 ubiquitin ligase RNF185 facilitates the cGAS-mediated innate immune response. Plos Pathog 13: e1006264. doi: 10.1371/journal.ppat.1006264
|
| [106] | Ni G, Konno H, Barber GN (2017) Ubiquitination of STING at lysine 224 controls IRF3 activation. Sci Immunol 2: In Press. |
| [107] |
Zhang J, Hu MM, Wang YY, et al. (2012) TRIM32 protein modulates type I interferon induction and cellular antiviral response by targeting MITA/STING protein for K63-linked ubiquitination. J Biol Chem 287: 28646–28655. doi: 10.1074/jbc.M112.362608
|
| [108] |
Tsuchida T, Zou J, Saitoh T, et al. (2010) The ubiquitin ligase TRIM56 regulates innate immune responses to intracellular double-stranded DNA. Immunity 33: 765–776. doi: 10.1016/j.immuni.2010.10.013
|
| [109] |
Wang J, Yang S, Liu L, et al. (2017) HTLV-1 Tax impairs K63-linked ubiquitination of STING to evade host innate immunity. Virus Res 232: 13–21. doi: 10.1016/j.virusres.2017.01.016
|
| [110] |
Liu Y, Li J, Chen J, et al. (2015) Hepatitis B virus polymerase disrupts K63-linked ubiquitination of STING to block innate cytosolic DNA-sensing pathways. J Virol 89: 2287–2300. doi: 10.1128/JVI.02760-14
|
| [111] |
Zhong B, Zhang L, Lei C, et al. (2009) The ubiquitin ligase RNF5 regulates antiviral responses by mediating degradation of the adaptor protein MITA. Immunity 30: 397–407. doi: 10.1016/j.immuni.2009.01.008
|
| [112] |
Wang Y, Lian Q, Yang B, et al. (2015) TRIM30α is a negative-feedback regulator of the intracellular DNA and DNA virus-triggered response by targeting STING. Plos Pathog 11: e1005012. doi: 10.1371/journal.ppat.1005012
|
| [113] |
Qin Y, Zhou MT, Hu MM, et al. (2014) RNF26 temporally regulates virus-triggered type I interferon induction by two distinct mechanisms. Plos Pathog 10: e1004358. doi: 10.1371/journal.ppat.1004358
|
| [114] |
Chen Y, Wang L, Jin J, et al. (2017) p38 inhibition provides anti-DNA virus immunity by regulation of USP21 phosphorylation and STING activation. J Exp Med 214: 991–1010. doi: 10.1084/jem.20161387
|
| [115] |
Lang X, Tang T, Jin T, et al. (2017) TRIM65-catalized ubiquitination is essential for MDA5-mediated antiviral innate immunity. J Exp Med 214: 459–473. doi: 10.1084/jem.20160592
|
| [116] | Liu B, Zhang M, Chu H, et al. (2017) The ubiquitin E3 ligase TRIM31 promotes aggregation and activation of the signaling adaptor MAVS through Lys63-linked polyubiquitination. Nat Immunol 18: 214–224. |
| [117] |
Narayan K, Waggoner L, Pham ST, et al. (2014) TRIM13 is a negative regulator of MDA5-mediated type I interferon production. J Virol 88: 10748–10757. doi: 10.1128/JVI.02593-13
|
| [118] |
Gao D, Yang YK, Wang RP, et al. (2009) REUL is a novel E3 ubiquitin ligase and stimulator of retinoic-acid-inducible gene-I. Plos One 4: e5760. doi: 10.1371/journal.pone.0005760
|
| [119] |
Oshiumi H, Matsumoto M, Hatakeyama S, et al. (2009) Riplet/RNF135, a RING-finger protein, ubiquitinates RIG-I to promote interferon-beta induction during the early phase of viral infection. J Biol Chem 284: 807–817. doi: 10.1074/jbc.M804259200
|
| [120] |
Gack MU, Shin YC, Joo CH, et al. (2007) TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature 446: 916–920. doi: 10.1038/nature05732
|
| [121] |
Jiang J, Li J, Fan W, et al. (2016) Robust Lys63-linked ubiquitination of RIG-I promotes cytokine eruption in early influenza B virus infection. J Virol 90: 6263–6275. doi: 10.1128/JVI.00549-16
|
| [122] |
Yan J, Li Q, Mao AP, et al. (2014) TRIM4 modulates type I interferon induction and cellular antiviral response by targeting RIG-I for K63-linked ubiquitination. J Mol Cell Biol 6: 154–163. doi: 10.1093/jmcb/mju005
|
| [123] |
Wang W, Jiang M, Liu S, et al. (2016) RNF122 suppresses antiviral type I interferon production by targeting RIG-I CARDs to mediate RIG-I degradation. Proc Natl Acad Sci USA 113: 9581–9586. doi: 10.1073/pnas.1604277113
|
| [124] |
Arimoto K, Takahashi H, Hishiki T, et al. (2007) Negative regulation of the RIG-I signaling by the ubiquitin ligase RNF125. Proc Natl Acad Sci USA 104: 7500–7505. doi: 10.1073/pnas.0611551104
|
| [125] |
Ning S, Campos AD, Darnay B, et al. (2008) TRAF6 and the three C-terminal lysine sites on IRF7 are required for its ubiquitination-mediated activation by the tumor necrosis factor receptor family member latent membrane protein 1. Mol Cell Biol 28: 6536–6546. doi: 10.1128/MCB.00785-08
|
| [126] |
Ning S, Pagano J (2010) The A20 deubiquitinase activity negatively regulates LMP1 activation of IRF7. J Virol 84: 6130–6138. doi: 10.1128/JVI.00364-10
|
| [127] |
Iwai K, Fujita H, Sasaki Y (2014) Linear ubiquitin chains: NF-kappaB signalling, cell death and beyond. Nat Rev Mol Cell Bio 15: 503–508. doi: 10.1038/nrm3836
|
| [128] |
Rieser E, Cordier SM, Walczak H (2013) Linear ubiquitination: a newly discovered regulator of cell signalling. Trends Biochem Sci 38: 94–102. doi: 10.1016/j.tibs.2012.11.007
|
| [129] |
Tokunaga F (2013) Linear ubiquitination-mediated NF-kappaB regulation and its related disorders. J Biochem 154: 313–323. doi: 10.1093/jb/mvt079
|
| [130] |
Tokunaga F, Iwai K (2012) Linear ubiquitination: a novel NF-kappaB regulatory mechanism for inflammatory and immune responses by the LUBAC ubiquitin ligase complex. Endocr J 59: 641–652. doi: 10.1507/endocrj.EJ12-0148
|
| [131] |
Shimizu Y, Taraborrelli L, Walczak H (2015) Linear ubiquitination in immunity. Immunol Rev 266: 190–207. doi: 10.1111/imr.12309
|
| [132] |
Ikeda F (2015) Linear ubiquitination signals in adaptive immune responses. Immunol Rev 266: 222–236. doi: 10.1111/imr.12300
|
| [133] |
Ikeda F, Deribe YL, Skanland SS, et al. (2011) SHARPIN forms a linear ubiquitin ligase complex regulating NF-kappaB activity and apoptosis. Nature 471: 637–641. doi: 10.1038/nature09814
|
| [134] |
Tokunaga F, Nakagawa T, Nakahara M, et al. (2011) SHARPIN is a component of the NF-kappaB-activating linear ubiquitin chain assembly complex. Nature 471: 633–636. doi: 10.1038/nature09815
|
| [135] |
Tian Y, Zhang Y, Zhong B, et al. (2007) RBCK1 negatively regulates tumor necrosis factor- and interleukin-1-triggered NF-kappaB activation by targeting TAB2/3 for degradation. J Biol Chem 282: 16776–16782. doi: 10.1074/jbc.M701913200
|
| [136] |
Niu J, Shi Y, Iwai K, et al. (2011) LUBAC regulates NF-kappaB activation upon genotoxic stress by promoting linear ubiquitination of NEMO. EMBO J 30: 3741–3753. doi: 10.1038/emboj.2011.264
|
| [137] |
Hostager BS, Kashiwada M, Colgan JD, et al. (2011) HOIL-1L interacting protein (HOIP) is essential for CD40 signaling. Plos One 6: e23061. doi: 10.1371/journal.pone.0023061
|
| [138] |
Zak DE, Schmitz F, Gold ES, et al. (2011) Systems analysis identifies an essential role for SHANK-associated RH domain-interacting protein (SHARPIN) in macrophage Toll-like receptor 2 (TLR2) responses. Proc Natl Acad Sci USA 108: 11536–11541. doi: 10.1073/pnas.1107577108
|
| [139] |
Rodgers MA, Bowman JW, Fujita H, et al. (2014) The linear ubiquitin assembly complex (LUBAC) is essential for NLRP3 inflammasome activation. J Exp Med 211: 1333–1347. doi: 10.1084/jem.20132486
|
| [140] |
Kirisako T, Kamei K, Murata S, et al. (2006) A ubiquitin ligase complex assembles linear polyubiquitin chains. EMBO J 25: 4877–4887. doi: 10.1038/sj.emboj.7601360
|
| [141] |
Emmerich CH, Schmukle AC, Walczak H (2011) The emerging role of linear ubiquitination in cell signaling. Sci Signal 4: re5. doi: 10.1126/scisignal.2001798
|
| [142] |
Tokunaga F, Sakata Si, Saeki Y, et al. (2009) Involvement of linear polyubiquitylation of NEMO in NF-kappa B activation. Nat Cell Biol 11: 123–132. doi: 10.1038/ncb1821
|
| [143] |
Inn KS, Gack MU, Tokunaga F, et al. (2011) Linear ubiquitin assembly complex negatively regulates RIG-I- and TRIM25-mediated type I interferon induction. Mol Cell 41: 354–365. doi: 10.1016/j.molcel.2010.12.029
|
| [144] |
Zhang M, Tian Y, Wang RP, et al. (2008) Negative feedback regulation of cellular antiviral signaling by RBCK1-mediated degradation of IRF3. Cell Res 18: 1096–1104. doi: 10.1038/cr.2008.277
|
| [145] |
Belgnaoui SM, Paz S, Samuel S, et al. (2012) Linear ubiquitination of NEMO negatively regulates the interferon antiviral response through disruption of the MAVS-TRAF3 complex. Cell Host Microbe 12: 211–222. doi: 10.1016/j.chom.2012.06.009
|
| [146] | Wang L, Wang Y, Zhao J, et al. (2017) LUBAC modulates LMP1 activation of NFκB and IRF7. J Virol 91: e1138–e1116. |
| [147] |
Orzalli MH, DeLuca NA, Knipe DM (2012) Nuclear IFI16 induction of IRF-3 signaling during herpesviral infection and degradation of IFI16 by the viral ICP0 protein. Proc Natl Acad Sci USA 109: E3008–E3017. doi: 10.1073/pnas.1211302109
|
| [148] |
Li T, Chen J, Cristea IM (2013) Human cytomegalovirus tegument protein pUL83 inhibits IFI16-mediated DNA sensing for immune evasion. Cell Host Microbe 14: 591–599. doi: 10.1016/j.chom.2013.10.007
|
| [149] |
Yu Y, Wang SE, Hayward GS (2005) The KSHV immediate-early transcription factor RTA encodes ubiquitin E3 ligase activity that targets IRF7 for proteosome-mediated degradation. Immunity 22: 59–70. doi: 10.1016/j.immuni.2004.11.011
|
| [150] |
van Gent M, Braem SGE, de Jong A, et al. (2014) Epstein-Barr virus large tegument protein BPLF1 contributes to innate immune evasion through interference with Toll-like receptor signaling. Plos Pathog 10: e1003960. doi: 10.1371/journal.ppat.1003960
|
| [151] |
Hu MM, Yang Q, Xie XQ, et al. (2016) Sumoylation promotes the stability of the DNA sensor cGAS and the adaptor STING to regulate the kinetics of response to DNA virus. Immunity 45: 555–569. doi: 10.1016/j.immuni.2016.08.014
|
| [152] |
Liang Q, Deng H, Li X, et al. (2011) Tripartite motif-containing protein 28 is a small ubiquitin-related modifier E3 ligase and negative regulator of IFN Regulatory Factor 7. J Immunol 187: 4754–4763. doi: 10.4049/jimmunol.1101704
|
| [153] |
Yang WL, Zhang X, Lin HK (2010) Emerging role of Lys-63 ubiquitination in protein kinase and phosphatase activation and cancer development. Oncogene 29: 4493–4503. doi: 10.1038/onc.2010.190
|
| [154] |
Yang Y, Kelly P, Schmitz R, et al. (2016) Targeting non-proteolytic protein ubiquitination for the treatment of diffuse large B cell lymphoma. Cancer Cell 29: 494–507. doi: 10.1016/j.ccell.2016.03.006
|
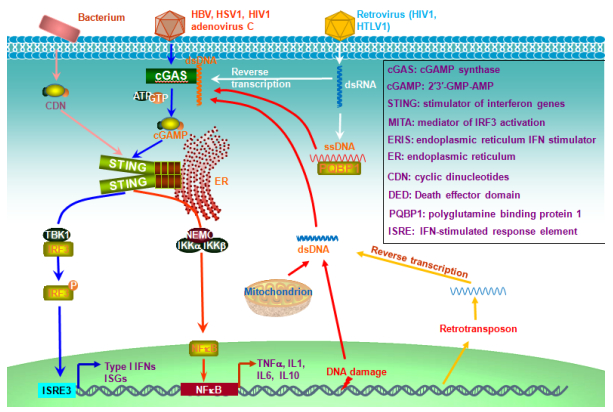
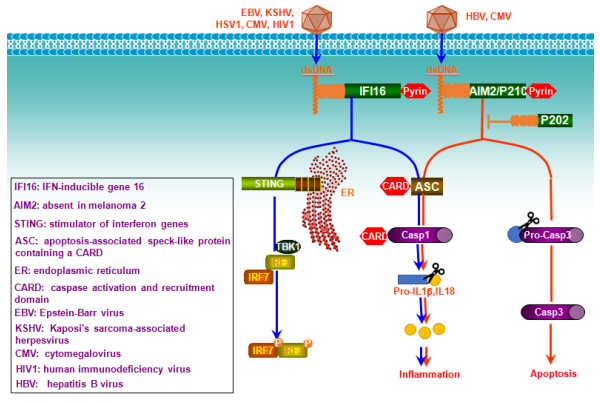
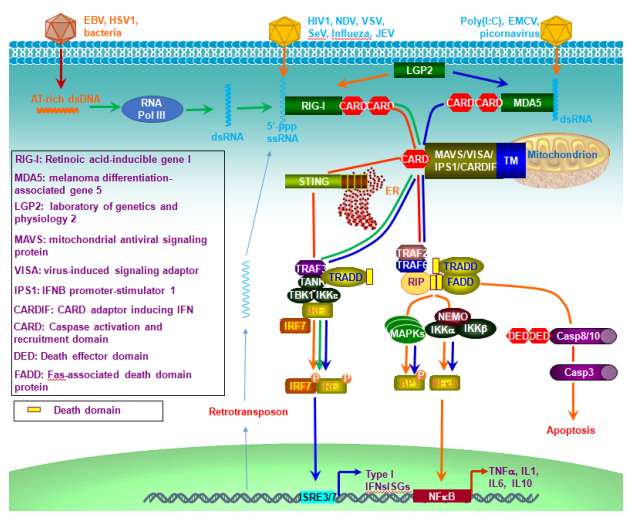
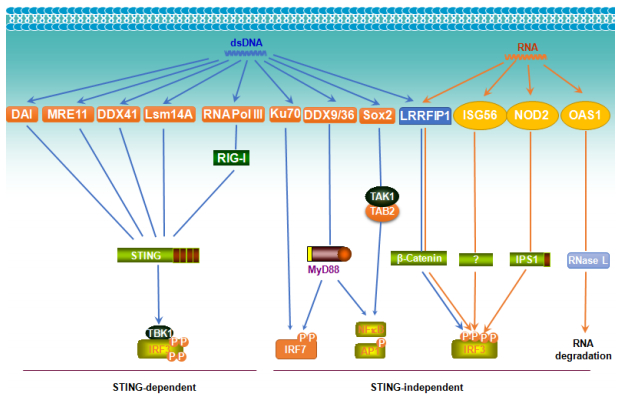
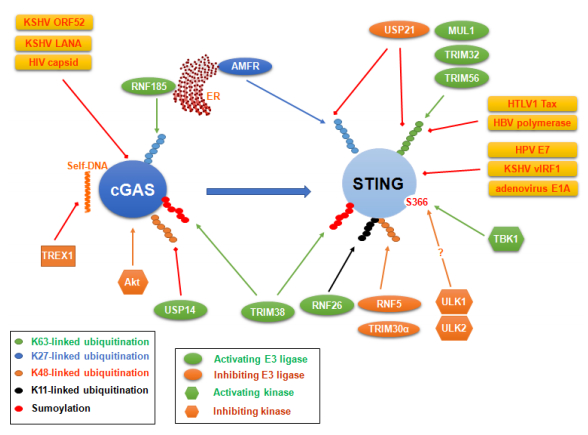
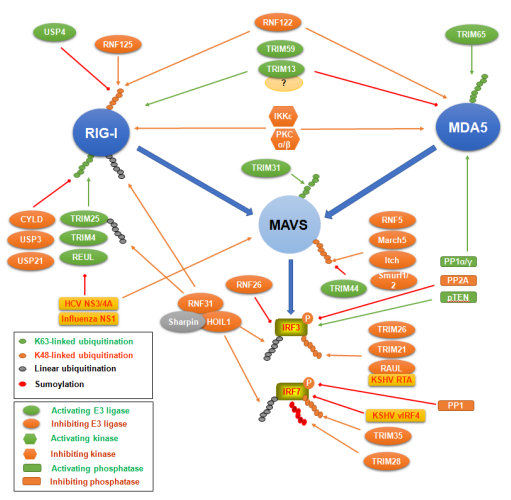
Figures(6)

Ling Wang, Shunbin Ning. “Toll-free” pathways for production of type I interferons[J]. AIMS Allergy and Immunology, 2017, 1(3): 143-163. doi: 10.3934/Allergy.2017.3.143







 DownLoad:
DownLoad: